Seabirds: strategic ornithological compensatory measures: review
A review of potential strategic ornithological compensatory measures, assessing their practical and ecological feasibility in the Scottish context
5 Ecological feasibility reviews
5.1 Ecological feasibility: Sandeel fishery closure
5.1.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, the Sandeel fishery closure has a low to medium effect on at least one demographic parameter of Kittiwake. We conclude that Sandeel fishery closures would have benefits to Kittiwake populations. However, the size of this benefit will most likely be small and will not be possible to quantify with high confidence. This conservation action would be best considered as a resilience-building measure that may assist Kittiwake in coping with additional pressures, such as climate change.
Note: This review was prepared prior to the announcement of a closure of the Sandeel fishery in Scottish Waters made in early 2024 (see note at the start of the corresponding practical feasibility review: Practical feasibility: Closure and management of forage fisheries (Sandeel, Herring, and Sprat) ).
5.1.2 Introduction and background
Many piscivorous seabird species are dependent on one or a few key prey fish species, often termed ‘forage fish’, due to their importance within marine ecosystems (Cury et al., 2011; Tasker and Sydeman, 2023). These same forage fish are also the target of industrial fisheries. As such there is potential for seabirds and fisheries to be competing for the same resource, with fisheries reducing prey availability to seabirds. However, these ecosystems are complex with both top-down and bottom-up ecological processes acting, often at the same time (Cury et al., 2000; Hunt Jr and McKinnell, 2006; Dickey-Collas et al., 2014).
Cury et al. (2011) reviewed the global impact of prey abundance on seabirds, establishing a general principle stating that seabird populations will suffer lower productivity where forage fish stocks are below one-third of maximum observed long-term biomass. Saraux et al. (2021) subsequently identified a lower threshold (15–18% of historical maximum biomass) below which seabirds start to exert top-down regulation on forage fish populations.
In the North Sea, Sandeels (fish belonging to the family Ammodytidae) are a key forage fish for multiple seabird species including the focal species for this review, the Black-legged Kittiwake (Harris and Wanless, 1997a; Dickey-Collas et al., 2014; Wanless et al., 2018). Due to their foraging ecology (surface feeding), Kittiwake are considered to be one of the most vulnerable seabird species to reduced Sandeel abundance (Furness and Tasker, 2000). However, the North Sea is a complex system with various not fully understood feedbacks operating between different parts of the ecosystem, which means that it is not straightforward to predict the consequences of alternative management options, particularly in the face of climate change (Engelhard et al., 2013; Dickey-Collas et al., 2014).
Sandeels, especially Lesser Sandeels (Ammodytes marinus), are the primary food source of Black-legged Kittiwake in the UK during the breeding season and are also the main target for the largest single species fishery in the North Sea. Sandeels are also considered to be particularly at risk from rising sea temperatures due to their specialised habitat requirements and limited ability to shift their distribution amid adverse conditions (Wright, 2020; Henriksen et al., 2021), and their abundance and quality have deteriorated over the past 30 years (Furness, 2007; Wanless et al., 2018). In the North Sea, Kittiwake breeding success and survival have been strongly linked to Sandeel availability (Harris and Wanless, 1997a; Furness and Tasker, 2000; Daunt et al., 2008; Searle et al., 2023a), such that years with higher Sandeel biomass were positively associated with Kittiwake productivity (Carroll et al., 2017). Furthermore, Kittiwake breeding success has had a consistent negative relationship with fishery pressure (Searle et al., 2023a), represented by the interaction between Sandeel population size and the proportion of the fish population harvested (Cook et al., 2014). In Shetland, for example, the breeding success of several seabirds, including Kittiwake, decreased substantially after the collapse of the Shetland Sandeel stock (Furness and Tasker, 2000). This highlights the crucial role that Sandeel populations play on the survival and population dynamics of Kittiwake, a declining and already vulnerable species.
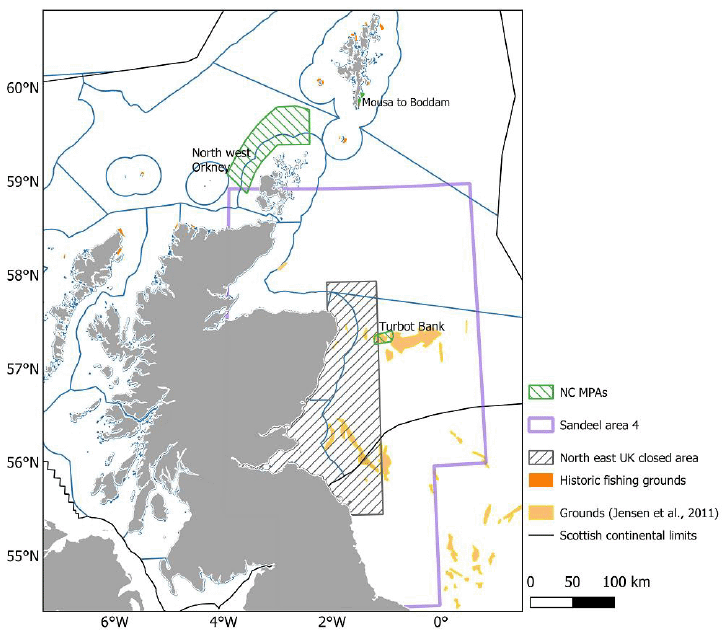
In 1990, an industrial Sandeel fishery began on the sandbanks of the Wee Bankie, Marr Bank, and Scalp Bank, 40 km off the southeast of Scotland. The fishery operated at the same time as the Kittiwake’ breeding season and had a considerable spatial overlap with Kittiwake foraging areas for colonies along the east coast of Scotland. Due to considerable concern that the fishery was adversely affecting seabird populations through direct competition, an area of approximately 21,000 km2 was closed to large scale fishing in 2000 (Figure 6) (Camphuysen, 2005; Wright, 2020). The closure was reviewed in 2007 leading to a recommendation to maintain the closure (STECF/SGMOS, 2007). Over 20 years have passed since the closure, and this is in a region where Kittiwake populations have been studied in detail before and since the closure (Daunt et al., 2008; Searle et al., 2023a). Therefore, there has been considerable research done to investigate the effect that the closure has had on Kittiwake populations.
5.1.3 Methods
The corresponding research question to this conservation action is: ‘Would Kittiwake populations benefit from widening the spatial extent of the Sandeel fishery closure?’
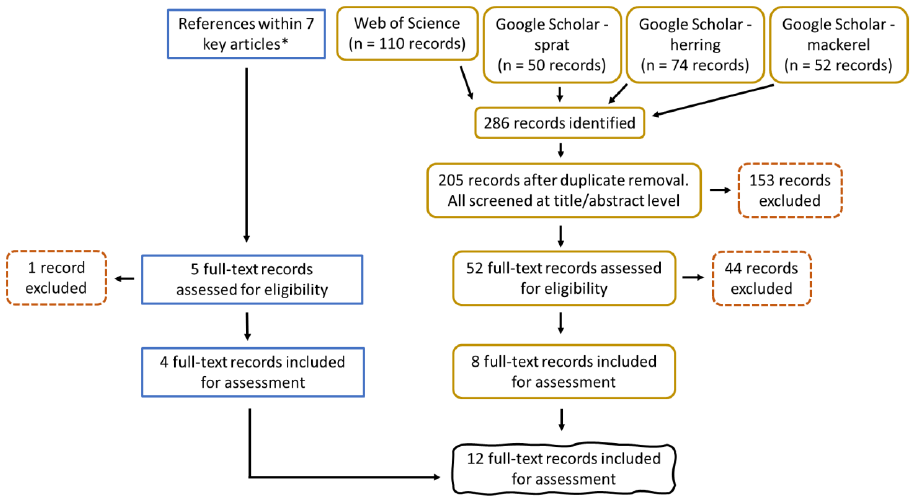
A detailed description of the research question, keywords, search strings, and study selection (Figure 7) is provided in section 2 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 18 references were included for the final review.
5.1.4 Results
5.1.4.1 General observations
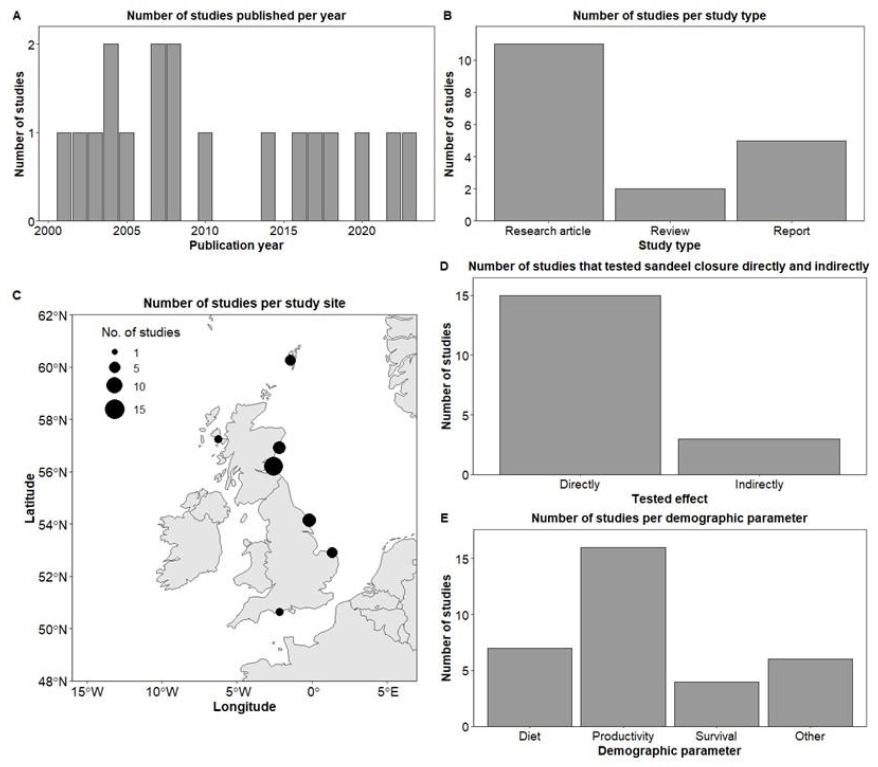
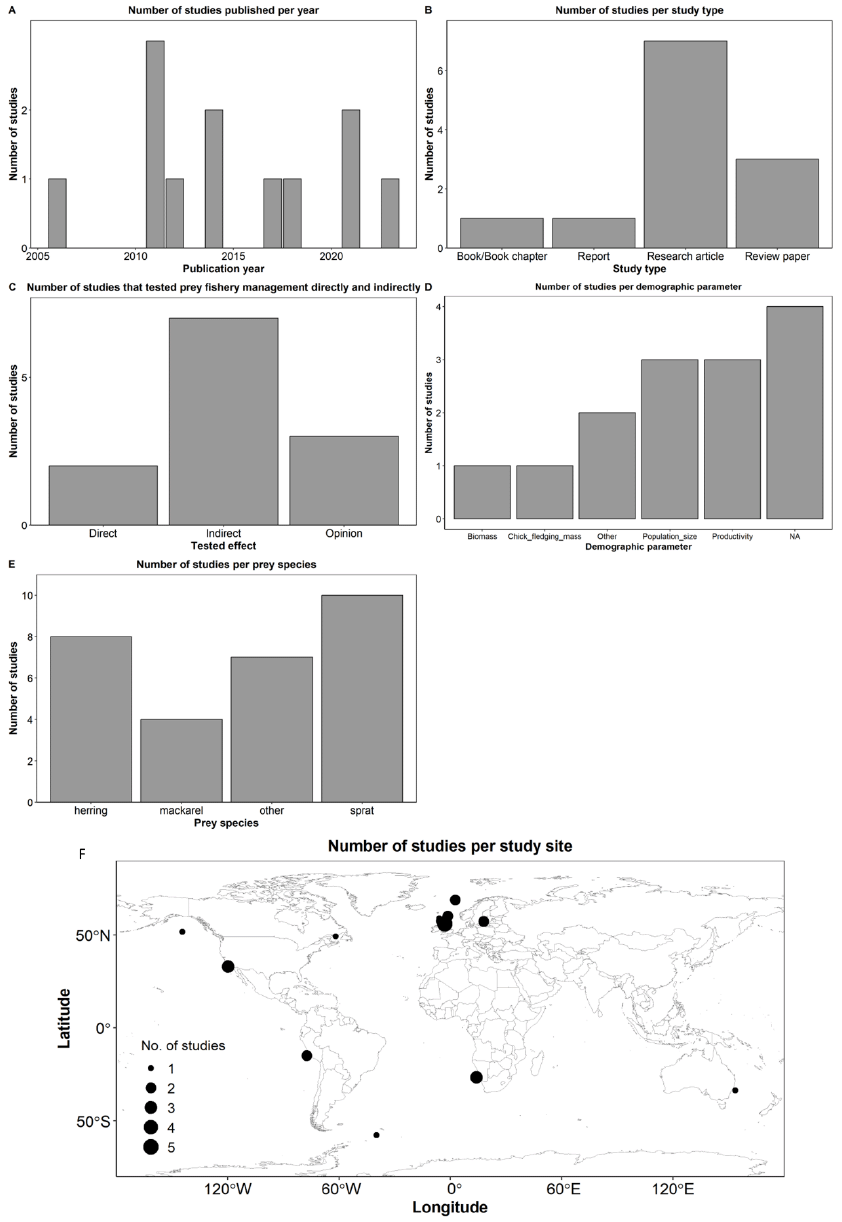
We identified 18 references, all published in the English language, that met our criteria of testing whether the Sandeel fishery closure had an effect on Kittiwake populations. The earliest publication resulting from this literature review was in 2001, and the latest in 2023 (Figure 8A). The median publication date is 2008, and although each year consistently generated at least one article, there is a gap between 2009 and 2013, for which we found only one study that passed our inclusion criteria. From the 18 references, 61% were research articles, 28% were reports, and 11% were review papers (Figure 8B). Most studies collected, used, or analysed data from the East Coast; 14 from Southeast Scotland (SES; mostly from the Isle of May), seven from Northeast England (NEE), six from Northeast Scotland (NES), four from East England (EE), four from Orkney and Shetland (OSS), two from Northwest Scotland (NWS), and two from Southwest England (SWE) (Figure 8C). Note that some studies used data from multiple regions. Out of the 18 studies, 15 (83%) tested the Sandeel fishery closure directly, while three (17%) tested it indirectly (Figure 8D). Most studies (89%) tested or analysed the impact the closure had on Kittiwake productivity or breeding success, 39% on diet, 22% on survival rates, and 28% on other parameters such as foraging movements, abundance, population size, and population trends (Figure 8E).
The most commonly used data collection and analyses methods were observational field data and diet sample collection of Kittiwake (most studies utilised information collected from the UK-wide Seabird Monitoring Programme; SMP), Sandeel stock assessments, modelling, and use of environmental variables. Data sets analysed ranged from one year of data collection to 42 years (median = 16 years). Among the authors, the one with the highest frequency authored eight references, whereas two other authors authored six records each.
5.1.4.2 Evidence scoring
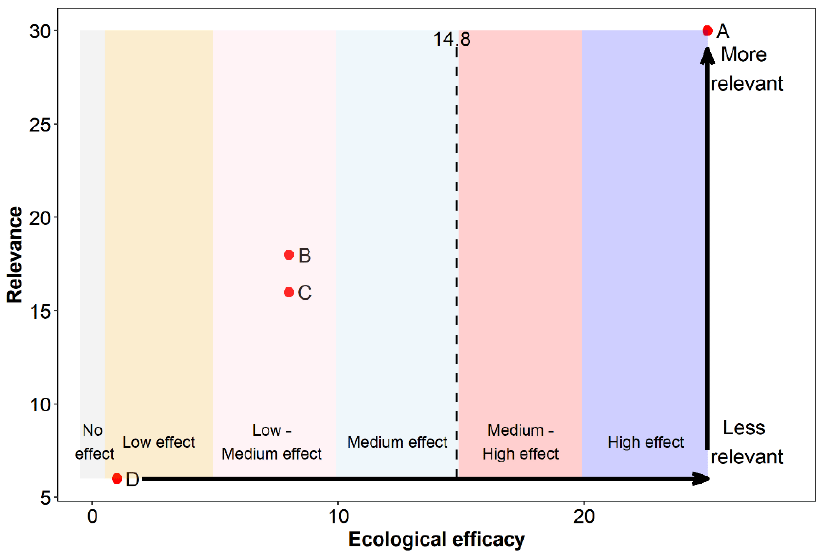
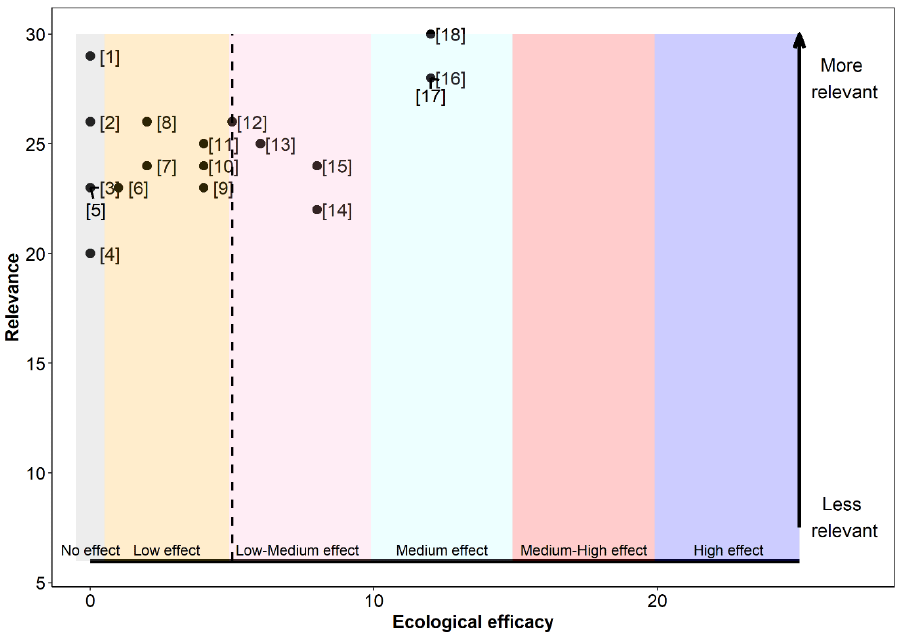
The relevance, as well as the ecological efficacy scores for all 18 references are shown in Table 17 and Figure 9 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Results from 72% of the assessed studies suggest that the Sandeel fishery closure had some sort of beneficial effect on at least one demographic aspect of Kittiwake, while the remaining 28% did not find it had any significant effect.
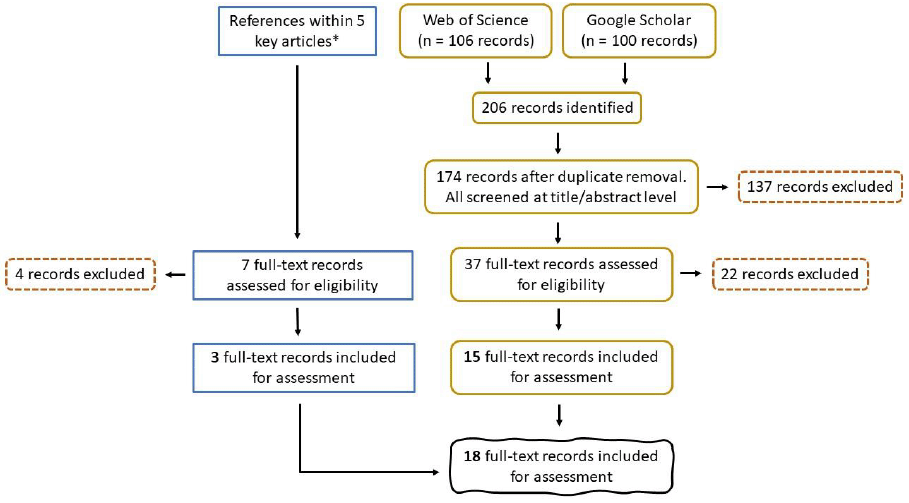
Table 17. Relevance and ecological efficacy scores, as well as the effect of the Sandeel fishery closure on Kittiwake for each assessed study. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of Sandeel fishery closure on Kittiwake, or those with low statistical inference; higher scores reflect studies that found a strong effect of closure on Kittiwake with high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
Overall, references were highly relevant for our assessment. Scores ranged between 20 and 30 out of a maximum possible score of 30, with a median and mean score of 24.5 and a mean average score of 25. For this reason, studies had a similar associated weight, ranging from 0.045 to 0.067 (Table 17).
The ecological efficacy, i.e. the likelihood that a Sandeel fishery closure is beneficial on at least one demographic parameter of Kittiwake, varied between studies. Out of a maximum possible score of 25, scores ranged between 0 (closure did not have an effect on at least one Kittiwake demographic parameter) and 12 (closure had a medium effect on Kittiwake). The median score is four, which indicates that half of the studies did not find a significant effect, or found a low effect, of the Sandeel closure on Kittiwake. The other half suggest that the Sandeel fishery closure had a low-medium or medium effect on Kittiwake.
The sum of all final weighted scores is five (i.e. the mean of ecological efficacy weighted by relevance), therefore, it can be concluded that the evidence overall support that a Sandeel fishery closure will benefit Kittiwake to a low-medium degree (Figure 9).
5.1.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this conservation action was eight out of the possible 15 (Table 18), suggesting there is a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of the Sandeel closure.
Table 18. Assessment of the confidence in the evidence for the Sandeel fishery closure.
This table can be viewed in Excel format.
5.1.4.4 Main findings: the effect of Sandeel fishery closure on Kittiwake
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, the Sandeel fishery closure has a low-medium effect on at least one demographic parameter of Kittiwake.
The key findings from the highest scored studies are summarised in Table 19. Overall, results from most, but not all, studies provide evidence that Kittiwake breeding success significantly increased after the Wee Bankie closure (Lewis et al., 2001b; Frederiksen et al., 2008; Ruffino et al., 2020; Searle et al., 2023a). Breeding success for Kittiwake on the Isle of May, for example, was substantially higher in the first three years of the closure (2000–2003) but had poor breeding success during subsequent years (such as 2004, 2007, and 2008). From 2009 onwards, there was a slight recovery, but it is unclear whether this improvement resulted from the Sandeel closure itself (Newell et al., 2016; Searle et al., 2023a) or from external factors. Changes in diet have also been recorded across time; there has been a decrease in 1+group (individuals at least one year old) Sandeel consumption, and an increase in sprat and 0-group (i.e. young of the year) Sandeel consumption (Lewis et al., 2001b; Wanless et al., 2018). Breeding success has strongly correlated with Sandeel total stock biomass; with breeding success lowest in years of low Sandeel abundance, and higher in years with moderate to high Sandeel stock biomass (Furness, 2007; Daunt et al., 2008). Years where Kittiwake had a higher proportion of Sandeel in their diet were associated with higher breeding success (Searle et al., 2023a). Lower breeding success, however, has also been significantly correlated with higher sea surface temperatures (Cook et al., 2014).
To support the recent consultation on options for spatial closures of English Waters to Sandeel fisheries (Defra, 2023a), Defra commissioned a review on benefits to seabirds that included new ecosystem modelling (Natural England et al., 2023). This report was not considered in our systematic review as it did not come up in our literature search likely due to its recent publication (March 2023). The report’s conclusions are in general agreement with our findings. The study concluded that there could be increased population resilience to seabird populations following a Sandeel fishery closure. There were some caveats around these findings with the seabirds modelled as a single group rather than individual species, so benefits to Kittiwake could not be specifically examined. Additionally, the authors noted that they had assumed constant conditions so do not consider future anticipated ecosystem changes driven by climate change.
The most recent and complete study to have analysed long-term Kittiwake productivity data and relate this to the Wee Bankie Sandeel fishery closure is Searle et al. (2023a). This study analysed data collected on the Isle of May (i.e. the same seabird colony used by most of the studies considered in this review) as well as data collected for other colonies of the East coast of the UK from 1986 to 2018. Breeding success after the fishery closure varied between colonies within the closure zone, ranging from -4% to 25%. Colonies such as Dunbar Coast and Farne Islands showed a continued decline since the closure, while the colony on the Isle of May had a 17% breeding success increase. Colonies within a control zone (i.e. outwith the Sandeel closure area), on the other hand, had a slight but non-significant decline in breeding success during that same time period. On the Isle of May evidence suggests a long-term decline in the overall importance of Sandeels in the diet of chicks, an increase in sprat and herring, and a shift from 1-group Sandeels to 0-group. The closure, however, did not restore the importance of Sandeels in the diet.
Numerous studies concur that Kittiwake populations are regulated by a variety of factors, including environmental conditions, fisheries, top-down processes, and Sandeel body condition. Consequently, disentangling and comprehending the individual effects of these factors can be difficult and challenging to tease apart (Lewis et al., 2001b; Wanless et al., 2007; Ruffino et al., 2020; Searle et al., 2023a).
5.1.4.5 Biases or Conflict of interest
For most of the studies considered the authors did not disclose any potential for biases or conflicts of interest. However, two of the studies were fully or partly funded by either a fisheries industry body (Furness, 2002) or by an offshore wind developer (Searle et al., 2023a). Note that we do not make judgement on whether these lead to an actual conflict of interest or influence study findings but provide the information for the readers’ awareness.
Table 19. Summary findings of the seven most relevant studies on the effect of Sandeel fishery closure on Kittiwake, in order of decreasing relevance.
This table can be viewed in Excel format.
5.1.5 Discussion
It is clear that seeking to manage the North Sea ecosystem for increased Sandeel availability would help maintain Kittiwake populations (Harris and Wanless, 1997a; Furness and Tasker, 2000; Daunt et al., 2008; Searle et al., 2023a). However, the evidence that further Sandeel fishery closures (i.e. beyond the existing closure of the Wee Bankie fishery off southeast Scotland) will deliver significant population level benefits to Kittiwake, required for this conservation action to be considered an ecologically feasible compensatory measure, is relatively weak.
The North Sea is a very well-studied ecosystem yet due to its complexity we do not have a sufficiently full understanding to confidently predict how changes in one component of the ecosystem affect others (Engelhard et al., 2013; Dickey-Collas et al., 2014; Régnier et al., 2019). More research might help improve our predictive power. However, this is likely ultimately limited with this being a complex system (Glaser et al., 2014) and one that is undergoing continuous longer term changes due to climate change, which is already leading to changes in the structure and functioning of the ecosystem (MacDonald et al., 2015; Mitchell et al., 2020; Olin et al., 2022).
A further fishery closure, in addition to the existing NE UK Sandeel closure area, may bring some ecosystem benefit through increasing ecosystem resilience, reducing the risk of a Sandeel stock collapse (e.g. Essington et al. (2015)), and by reducing prey competition with seabirds, particularly in areas near to breeding colonies where fisheries could have the most significant impact on breeding birds (Pichegru et al., 2010; Sherley et al., 2015; Hentati-Sundberg et al., 2021b). The spatial component of prey availability and how this could be impacted by fisheries may be particularly relevant for Sandeel which have low dispersal distances (Wright et al., 2019), presumably increasing potential for local depletion.
The most comprehensive study to date on the effect of the existing Sandeel fishery closure, Searle et al. (2023a), generally found increases in breeding success for Kittiwake colonies within the closure region but slight declines (though non-significant) outwith this zone. However, despite the increases in breeding success, the Kittiwake populations have not recovered to pre-fishery levels even 20 years post-closure. The authors considered the implications of their findings for use of fisheries management as a strategic conservation tool. They noted that their models generally explained relatively low proportions of variance (4–50%) and that the system is impacted by environmental changes, thus caution should be applied if trying to produce quantitative predictions on the benefits from fishery closures into the future. They also noted that studies to monitor the effects of such management interventions will generally take many years, potentially decades, to provide clear conclusions which would limit the potential for adaptive management.
The impacts of Sandeel fisheries on Kittiwake (and other seabird populations) in recent years are likely relatively less than would have been detected in earlier decades due to overall reduced levels of fishing effort and the size of climate change effects. Overall fishing mortality on Sandeel in the North Sea is now <20% of overall annual mortality, which is historically low, and considered to be at a low exploitation rate (the ratio of fisheries mortality to natural mortality) (Engelhard et al., 2013). Over the past few decades, the zooplankton community has changed substantially, particularly along the east coast of Scotland, leading to substantial long-term declines in total energy available to Sandeel, particularly to 0-group Sandeel (Olin et al., 2022). Together these changes are affecting both Sandeel abundance and likely availability (a function of the spatio-temporal abundance) to Kittiwake with potential for a trophic mismatch (Scott et al., 2006; Burthe et al., 2012; Régnier et al., 2019; Mitchell et al., 2020).
5.1.5.1 Knowledge gaps and future directions
If Sandeel fisheries are to continue operating, then it will be crucial that there is better understanding of the spatial and temporal scale of fishery effects at scales relevant to Kittiwake. This could be achieved through a mixture of model-based studies (e.g. Searle et al. (2018); Hentati-Sundberg et al. (2021b)) and new data collection. Fish surveys that more directly measure prey availability and quality for seabirds are crucial, as most existing studies make inferences using fisheries monitoring data that are rarely optimal. For example, surveys are often conducted in a different season or at a different spatial scale than that at which seabird-prey interactions occur; Sandeel assessments are primarily based on dredge sampling during winter which is not directly representative of availability in the water column during the Kittiwake breeding period. Therefore, there is a need for whole-ecosystem models that are developed and parameterised to match the scales at which seabird-prey interactions occur (see e.g. Smith et al. (2011)).
Most studies of Kittiwake and prey in the UK have focussed on North Sea populations, especially from the long-running study on the Isle of May. These have delivered valuable insights into the complex system but there is increased uncertainty when extrapolating out to other parts of Scotland and the wider UK. With the recent completion of the national seabird census (‘Seabirds Count’) (Burnell et al., 2023) new up-to-date population data is now available for Kittiwake populations within Scotland, which could allow studies of regional differences including re-running earlier analyses with longer time-series and/or at wider spatial scales (e.g. Cook et al. (2014); Carroll et al. (2017)).
Most studies to date looked at Kittiwake productivity with relatively few relating variation or changes in survival to fisheries and prey (Figure 8). Given that a small increase in adult survival will, all else being equal, lead to a greater population level effect than a similarly small increase in productivity, then this should be investigated. Through the Seabird Monitoring Programme, adult survival (or more accurately resighting) data are available for a number of colonies, however this is still relatively few so ideally such long-term field studies would be increased and expanded (see O’Hanlon et al. (2021)).
The wider Sandeel fishery closures announced by the UK and Scottish Governments in early 2024[2] will potentially provide an opportunity to undertake similar Before-After-Control-Impact Sandeel closure analyses to those that have been undertaken for the southeast of Scotland. While these would be of less direct relevance to Scotland, these would provide a form of study replication while also improving our understanding by increasing the range of ecological conditions represented which should enhance predictive power.
5.1.6 Conclusion
Overall, we conclude that Sandeel fisheries closures would have benefits to Kittiwake populations, however the size of this benefit is generally small and is not possible to quantify with higher confidence. This conservation action is thus best considered as a resilience-building measure that may assist Kittiwake in coping with additional pressures, such as climate change. This conclusion aligns with the findings of Natural England et al. (2023). The evidence reviewed suggests that Sandeel abundance and availability is largely driven by processes other than fisheries including climate and piscivorous fish species (Engelhard et al., 2013; Dickey-Collas et al., 2014). Therefore, we have limited confidence that further closures could deliver an ecologically meaningful gain in Sandeel populations with ensuing benefits in Kittiwake and consequently, it cannot be concluded that such closures would represent an effective conservation action in terms of a compensatory measure. However, as Montevecchi (2023) noted it is also true that the only factor potentially determining prey availability to seabirds that we can generally control directly is fisheries.
5.2 Ecological feasibility: Fishery closure or enhanced management of prey fisheries (Sprat, Herring, and Mackerel)
5.2.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, the enhanced management of fisheries targeting seabird prey fish species has a low to medium effect on at least one demographic parameter of seabirds, but this is highly dependent on the seabird species and the current fisheries management practice. Whether a population level benefit can be anticipated following changes in fisheries management for a seabird species will require careful consideration of the specific ecological context, and it will rarely be possible to confidently form quantitative predictions on the level of benefit.
5.2.2 Introduction and background
Several piscivorous seabird species rely on one or a few key prey fish species (Cury et al., 2011; Pikitch et al., 2014; Tasker and Sydeman, 2023). Coincidentally, these same forage fish are targeted by industrial fisheries. This present the potential for competition between seabirds and fisheries for the same resource, with fisheries reducing prey availability to seabirds. However, these ecosystems are complex with both top-down and bottom-up ecological processes acting, often at the same time (Cury et al., 2000; Hunt Jr and McKinnell, 2006; Fauchald et al., 2011; Dickey-Collas et al., 2014).
Cury et al. (2011) estimated that seabird populations will suffer lower productivity where forage fish stocks are below one-third of the maximum observed long-term biomass. Saraux et al. (2021) subsequently identified a lower threshold (15–18% of historical maximum biomass) below which seabirds start to exert top-down regulation on forage fish populations (above this threshold predation by seabirds represents only a small proportion of overall fish mortality). However, in another global meta-analysis, marine predator productivity, including piscivorous fish, birds, and mammals, was found to be only rarely impacted by prey abundance (Free et al., 2021).
A global analysis of seabird distributions, considering seabird prey consumption and fisheries activity suggests that NW Europe, particularly around the UK, is an area of relatively high overlap in resource demands between seabirds and fisheries, and as such there is a higher expectation of potential resource competition (Karpouzi et al., 2007). There are multiple examples of seabird-prey systems in NW Europe where fisheries are considered to have had a role in reducing the availability of prey fish to seabirds leading ultimately to population level impacts. For example, a collapse in the Norwegian Herring stock was partly implicated in a decrease from 1.4 million to <600 thousands pairs of Atlantic Puffin at Røst, an island in Northern Norway, between the late 1970s and late 1990s. Around the UK the best studied system is of the dependence of seabirds in the North Sea on Sandeel and consequent impacts from reduce prey abundance (see also Ecological feasibility: Sandeel fishery closure). The breeding success of several seabirds, including Kittiwake, decreased substantially after the collapse of the Shetland Sandeel stock (Furness and Tasker, 2000). Kittiwake breeding success has had a consistent negative relationship with fishery pressure (Searle et al., 2023a), represented by the interaction between Sandeel population size and the proportion of the fish population harvested (Cook et al., 2014).
To understand which fisheries have potential to impact seabird populations in Scotland, we must first understand the diet of seabirds, then relate that to which prey items are also the target of fisheries. However, it should be noted that there is also the potential for indirect impacts from fisheries targeting other components of the food chain; as most of the prey species targeted by seabirds are forage fish (low trophic level), fisheries targeting predatory fish can alter the levels of natural predation on forage fish. Where natural predation (the majority of which is from predatory fish) is a large component of fish mortality, then management of fisheries targeting predatory fish has potential to have greater influence on the availability of prey to seabird population than changes in management of fisheries targeting the prey fish themselves (Österblom et al., 2006; Bakun et al., 2009; Reilly et al., 2014).
Our understanding of seabird diet in the UK is better for the breeding period than for the non-breeding season. The existing evidence on seabird diets in the UK was collated by the Marine Ecosystem Research Programme which is available as a publicly available database (via the seabirddietDB package in the R statistical environment, Krystalli et al. (2019)). Sandeel have been the key prey for many North Sea breeding seabirds, though their relative dietary importance has reduced over the past decades (Wanless et al., 2018). A study of Common Guillemot diets around the UK (Anderson et al., 2014) found great variation in chick diets depending on location, while Lesser Sandeel were the most frequent prey, clupeids (e.g. Sprat) and juvenile gadids were also important dietary components. Gannet take similar species but typically target larger fish, both of the same species (i.e. older age classes), and of others, particularly Mackerel in the North Sea (Lewis et al., 2003).
This chapter takes a broader approach than that focussing on Sandeel (see Ecological feasibility: Sandeel fishery closure) given the wider scope of multiple fisheries and multiple focal species. We therefore started by identifying key prey fisheries to focus the review on, largely following a previous study focussed around ecosystem based management of Scottish fisheries (Heath et al., 2017).
5.2.3 Methods
5.2.3.1 Prey fish species to include
The starting point for this review was to identify what seabird prey fish species would be relevant to consider in the review. To be included, the fish species need to be those that are prey fish of at least one focal seabird species but that are also targeted by fisheries in Scotland. Heath et al. (2017) reviewed evidence around interactions between commercial fisheries and natural predators (including seabirds) in Scotland. The review included consideration of potential for bycatch mortality, impacts on prey availability and potential for predator species to benefit from discards. However, only the sections pertinent to prey availability were considered for this review. They compiled a table classifying the likelihood for significant interactions between fisheries and predators (Table 2 in Heath et al. (2017)) which summarises the findings from a workshop (held in 2016 with a number of invited experts – see page 8 of Heath et al. (2017) for participant list). Below (Table 20) is an edited and re-arranged subset from this table including our focal seabird species and the conclusion of Heath et al. (2017) on the potential for fishery related prey depletion to impact a seabird species (‘predator likely to be affected by fishery’).
Table 20. Prey fish species identified by Heath et al. (2017) where there was potential for seabird species to be impacted by fisheries targeting these species. An ‘x’ denotes where a potential negative interaction is identified, a dot ‘.’ where no negative interaction was identified, and in one case a question mark ‘?’ where this was not clearly stated. After table 2 in Heath et al. (2017).
This table can be viewed in Excel format.
For the review we included all the prey fish species identified by Heath et al. (2017) where there was potential for at least one seabird species to be impacted by fisheries targeting those species. However, we excluded Sandeel, as these are the target of another review (see Ecological feasibility: Sandeel fishery closure), and rockling as on investigation these do not appear to be a commercially targeted fish species in Scottish Waters. The final list of prey fish species was also discussed with the project steering group (20th March 2023) where there was general agreement on the species to include, though suggestion to also consider gadoid species as the younger age classes of these are preyed on by seabirds. However, we decided to exclude these as they were not identified by our key source review (Heath et al., 2017) likely due to the abundance of the early age classes of these species being determined by complex ecosystem interactions and not primarily by fisheries targeting the adult age classes (see e.g. (Engelhard et al., 2013; Dickey-Collas et al., 2014)).
The final list of prey fish species included were Sprat, Herring, and Mackerel.
5.2.3.2 Research questions, keywords, search strings, and study selection
The corresponding research question to this conservation action is: ‘Would the focal seabird species benefit from enhanced management of fisheries that target seabird prey fish species in Scotland?’ In addition to the key question, we also sought to identify which prey species may be most impacted by fisheries and which seabird species could most benefit from any changes in management.
A detailed description of the research question, keywords, search strings, and study selection (Figure 10) is provided in section 3 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 12 references were included for the final review.
5.2.4 Results
5.2.4.1 General observations
We identified 12 references that met our criteria of measuring the effect of management of fisheries targeting seabird prey fish populations to affect seabird populations, all published in English. The literature review yielded publications ranging from 2006 to 2023 (Figure 11A). The median publication date is 2014. The years 2011, 2014, and 2021 had multiple publications. Most references were research articles (58%), but we also identified three review articles, one book chapter, and one commissioned report (Figure 11B).
Most studies, 58%, considered the impacts on seabird populations from fisheries management indirectly (Figure 11C). That is most studies did not include an experimental design approach (either using an experimental manipulation or opportunistically following changes in fishery management) where responses could be assessed before/during/after changes in fisheries management. Two studies did assess impacts directly, either via simulation or in an opportunistic study following seabird populations before/during/after changes in fisheries management. Finally, three studies were classified as opinion based which were in all cases narrative-based reviews thus did not include any quantitative or qualitative analysis of data. A variety of demographic related parameters were considered by the studies (Figure 11D), with productivity and population size the most common. Four studies did not clearly assess any specific demographic parameter (indicated by NA in Figure 11D); these were all review/report-based studies that had more general discussions on impacts on seabird populations that didn’t consider specific parameters (e.g. by instead assessing on a vulnerability scale). Many of the studies included multiple prey species (Figure 11E), of the three focal prey species in this review, sprat was most common (83%), followed by herring (67%), and mackerel (33%). Several studies included additional prey species (58%), these were mostly international studies or those including Sandeel which were excluded from this review given the focal review considering this fishery management (see Ecological feasibility: Sandeel fishery closure). One of the review inclusion criteria (Table A 2) was that studies included the North Atlantic or Europe, which is reflected in the geographical distribution of studies (Figure 11F), however as several studies included multiple locations/regions (particularly reviews) a number of sites beyond this were included.
Most studies combined multiple types of data collection, usually using fisheries-based statistics for prey fish, and colony observations for the seabirds studied. Analyses included more basic correlational studies and those using more sophisticated multivariate models. A few studies used ecosystem models either directly (i.e. bespoke for a given study) or indirectly (in review based studies where previous ecosystem modelling was considered). Three studies (Cury et al., 2011; Smith et al., 2011; Free et al., 2021) undertook analysis of multiple prey-predator systems (all including datasets from Scotland) seeking to come to more general results on the relationship between prey fish biomass and their predators (all including seabirds, though two also including other marine predators).
Though several of the studies included at least some of the same study systems (e.g. North Sea or California Current) only a few authors contributed to more than one of the 12 studies, with six authors appearing twice, all others only appeared once.
5.2.4.2 Evidence scoring
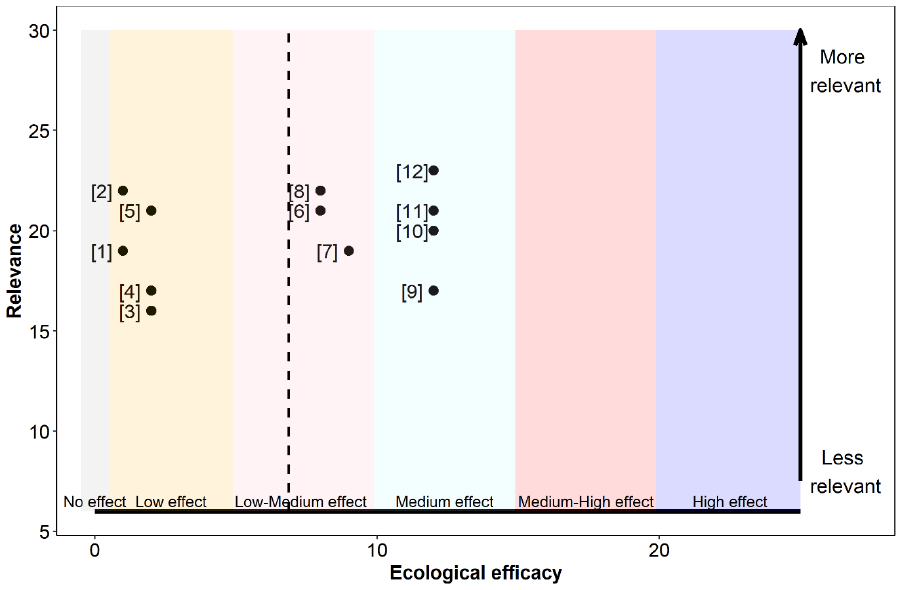
The relevance, as well as the ecological efficacy scores for all 12 references are shown in Table 21 and Figure 12 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). All studies suggested potential for some beneficial effect on at least one seabird species from management of fisheries targeting seabird prey fish species. However, the strength of an effect varied and, in several cases, while it was concluded that fisheries could impact prey availability to seabirds, this would only be significant if fishery activity were to increase.
The ecological efficacy, i.e. the likelihood that changes in the management of fisheries targeting seabird prey fish species is beneficial on at least one seabird demographic parameter, varied between studies. Out of a maximum possible score of 25, scores ranged between 1 and 12. The median score is 8.
For most of the studies included authors did not specifically compare between specific changes in management (reflected in all except one being assessed as providing non-direct evidence –Table 21). Rather most studies were focussed on either: identifying thresholds (e.g. ultimately equating to stock biomass but often expressed in relation to fisheries catch targets which for many species are currently based on maximum sustainable yield – MSY); on determining the relative balance of top-down (including predation, mostly from piscivorous fish, and fisheries removals) and bottom-up processes (for most prey species this equates proximally to zooplankton abundance and energy content which ultimately is primarily driven by primary productivity); or in reviewing previous studies. As such the ecological efficacy score here more reflects evidence that changes in fisheries management could affect seabird populations rather than more directly demonstrating a benefit.
The sum of all final weighted scores (i.e. the mean of ecological efficacy weighted by relevance) is 6.9. Therefore, it can be concluded that the evidence supports management of fisheries targeting seabird prey fish species being of benefit to seabirds, to a low-medium degree (Figure 12). However, as previously discussed, the potential for seabird populations to benefit will depend on the current fisheries management (i.e. to what extent fisheries pressure can be reduced with enhanced management).
Table 21. Relevance and ecological efficacy scores of each assessed study of enhanced management of fisheries targeting seabird prey fish species. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a strong effect of fisheries management on seabirds, or those with low statistical inference; higher scores reflect studies that found a strong effect of enhanced fisheries management with high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
5.2.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this conservation action was 10 out of the possible 15 (Table 22), suggesting that we have a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of enhanced management of fisheries targeting seabird prey fish species.
Table 22. Assessment of the confidence in the evidence of enhanced management of fisheries targeting seabird prey fish species.
This table can be viewed in Excel format.
5.2.4.4 Main findings: the effect of management of prey fisheries
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, the enhanced management of fisheries targeting seabird prey fish species has a low-medium effect on at least one demographic parameter of seabirds, but this is highly dependent on the seabird species and the current fisheries management practice (i.e. whether changes in current management would bring benefits to seabirds).
The key findings from the most relevant studies are summarised in Table 23. Overall, the studies all provided at least some support for fisheries management having potential to benefit seabird populations via driving changes in prey availability. Most studies considered fishery impacts (positive or negative depending on the cases studied) on seabirds indirectly via relating prey fish biomass to seabird demographic parameters. In these cases the effects of different fishery management targets (e.g. maximum sustainable yield) were considered indirectly by equating these to the associated fish biomass levels expected for a given fishery stock management targets. Most studies suggested that there was not a simple linear relationship between fish biomass and seabird demography, but rather that at a lower critical threshold of prey biomass, seabird population size would start to be significantly (in some cases dramatically) reduced.
Cury et al. (2011) studied 19 time-series (comprising 14 seabird species) where seabird productivity could be matched to prey fish biomass data. From this they identified a near universal threshold of prey fish biomass where, below one third of historical maximum biomass, productivity began to decline rapidly as biomass decreased further. This level of one third of historical maximum biomass generally occurs at a level of fisheries exploitation below the common fishery target of maximum sustainable yield (MSY). Guillemette et al. (2018) performed a similar analysis though focussed on a single prey-predator system of Gannet and Mackerel in the Gulf of St. Lawrence in Canada. They found a lower threshold of 8% of historical maximum biomass below which productivity declined rapidly. Gannet breeding success was on average 73% (i.e. 73% would successfully raise a chick to fledging each breeding season) when Mackerel biomass levels were greater than 8% of historical biomass but breeding success was only around 31% below this 8% threshold. Smith et al. (2011) undertook a similar analysis to Cury et al. (2011), though instead of time-series analysis they used ecosystem models. They identified that for many top marine predators, including seabirds, negative impacts on the marine predators (as measured in terms of their biomass) generally occurred at fishery exploitation rates below MSY. Free et al. (2021) undertook a similar analysis to (Cury et al., 2011) using time-series data of marine predators and prey, but rather than using productivity rates they used population count data to derive population growth rates (i.e. change in population size over time). For most seabird populations they did not find a relationship between prey fish biomass and seabird population growth rates. Overall, these studies suggested that seabirds will benefit from having fishery targets set slightly below MSY (with the exception of Free et al. (2021)).
The studies above were all focussed on wide spatial scales (whole ecosystems, e.g. North Sea) so did not account for local/regional spatial effects (e.g. potential for local prey depletion). Similar to the studies above, Hentati-Sundberg et al. (2021b) investigated the potential impacts of different fishery targets (on Herring and Sprat) on Guillemot and Razorbill. However, they took a different approach using a spatially explicit simulation model which could account for local prey depletion and travel costs for the seabirds as well as specific traits (e.g. diving depths). Their results were broadly in line with the studies above, finding that successful breeding could generally occur at relatively low biomass levels (equating to less than MSY), though they noted that these findings may be site-specific with potential for different results if modelling other locations (they focussed on a site in the Baltic Sea).
The only study included that directly related changes in fishery management to seabird population responses was Jennings et al. (2012). Their study focussed on Common Tern and Herring in the Firth of Forth, finding that the colony counts for the terns decreased significantly during the period when a fishery was active with a subsequent recovery when the fishery was closed. While this response was quite strong the contrast in prey levels were likely quite large; although Herring levels were not measured in the study it was noted that during the period the fishery was active Herring numbers collapsed.
Several studies took a more holistic approach, reviewing the relative contributions of top-down (both natural predation and fisheries) and bottom-up processes (mediated via zooplankton) in driving prey fish abundance and relating this to seabird populations. Fauchald et al. (2011) studied the North Sea ecosystem, relating seabird abundance during the non-breeding season (at sea abundance) to Herring and Sprat abundance and to zooplankton abundance. They found evidence for both top-down and bottom-up processes operating, which has been termed a wasp-waist system (Cury et al., 2000), as relatively few mid-trophic level species drive the abundance of both lower and higher trophic levels. Fauchald et al. (2011) related their results to fisheries management suggesting that their findings lend support to the potential for the harvest of prey fish by fisheries to affect both lower (zooplankton) and higher (seabirds and other predators) trophic levels. Two studies reviewed the current scientific knowledge of the North Sea ecosystem (Engelhard et al., 2013; Dickey-Collas et al., 2014). Together these found that prey abundance is highly variable and that this is largely driven by climate and natural predation (mostly from predatory fish). They also note potential for fisheries to impact on prey abundance and for this to impact on predators (including seabirds).
Table 23. Summary findings of the six most relevant studies on the effect of enhanced management of fisheries targeting seabird prey fish species, in order of decreasing relevance.
This table can be viewed in Excel format.
While the majority of studies focussed on prey abundance or overall biomass, Österblom et al. (2006) investigated the effects of both prey quality and quantity (in this case Sprat) on Guillemot as measured by chick fledging weights. They found that Sprat quality (weight-at-age) affected chick fledging-mass, but Sprat abundance did not affect fledging-mass. This result contrasts with most of the studies above, in that this study reported the potential for intermediate levels of fishing to increase Sprat quality (by reducing intra-specific competition) with positive impacts for chick fledging mass.
One study, Heath et al. (2017), was highly relevant as it focussed on identifying fisheries in Scotland where there was potential for ecosystem impacts, including specific consideration of seabirds and their prey species (where these were also target species of fisheries). However, the findings were based principally on expert opinion (via a workshop) rather than empirical study. As such this report sought to identify where there is potential for fisheries to impact prey availability to seabirds (the report covers other aspects of fisheries/ecosystem interactions but those were not relevant to this review) rather than to form firm conclusions on where additional fishery management would be beneficial.
Montevecchi (2023) provides an up-to-date review of global understanding on the potential for fisheries to impact prey availability to seabirds and thus of whether changes in fisheries management could benefit seabird populations (it also covers aspects of seabird/fisheries interactions not pertinent to our review). In contrast to the other three studies included here that have a global focus (Cury et al., 2011; Smith et al., 2011; Free et al., 2021), Montevecchi (2023) does not include any new data analysis nor collation but provides a narrative overview. Overall Montevecchi (2023) concludes that fisheries can impact prey availability to seabirds but that such fishery effects can be swamped by natural variability in prey populations making attributing effects from fisheries on seabird populations challenging.
5.2.4.5 Biases or Conflict of interest
Of the 12 studies, five included potentials for conflicts of interest (note we do not make judgement on whether these lead to an actual conflict of interest or influence study findings but provide the information for the readers’ awareness). Two studies were wholly or partly funded by a nature conservation organisation (Österblom et al., 2006; Hentati-Sundberg et al., 2021b), one part-funded by a fisheries sustainability accreditation organisation (Smith et al., 2011), one included an author funded by a (non-fishery) industry organisation (Jennings et al., 2012), and one an author supported by a fishery industry body (Free et al., 2021). The authors of the other studies did not disclose any potential biases or conflicts of interest.
5.2.5 Discussion
Overall, this review found evidence that the enhanced management of fisheries targeting seabird prey fish species can lead to population level benefits to seabird populations. However, this is highly dependent on the seabird species and the current fisheries management. Given the broad scope of this review considering multiple seabird species and prey fish, it was not possible to examine in detail individual species or fisheries (unlike for the review on Sandeel and Kittiwake, see Ecological feasibility: Sandeel fishery closure).
This review was focussed on fisheries targeting seabird prey fish, as such it does not consider all the ways in which fisheries can impact food availability to seabirds. Fisheries can provide direct food subsidies to seabirds via discards and offal provision (Garthe and Huppop, 1996; Votier et al., 2010), though changes in discarding regulations are thought to have reduced the availability of this (Bicknell et al., 2013; Sherley et al., 2020). While intuitively fisheries targeting prey fisheries are expected to have the greatest (fishery) impact on prey availability to seabirds, fisheries of predatory fish can also affect the abundance of prey fish by changing the ecosystem balance and changing levels of natural predation (Furness, 2002; Kadin et al., 2019). Changes in the overall balance between top-down processes (such as predation by predatory fish) and bottom-up processes (e.g. zooplankton abundance) can impact the abundance of low trophic level fish populations (termed forage fish), with fisheries targeting predatory fish having potential to lead to switching between different ecosystem states (termed regime shifts) (Österblom et al., 2006; Bakun et al., 2009). Additionally fisheries can alter fish habitat by the action of the fishing gear leading to physical disturbance to seafloor habitats (Rijnsdorp et al., 2020) (see also: Manage supporting habitats that relate to prey availability for seabirds). While these general principles are understood, these ecosystems have complex interactions which limit our capacity to predict ecological outcomes, including for the well-studied North Sea (Engelhard et al., 2013; Dickey-Collas et al., 2014).
The impacts of prey abundance on seabird populations have been well studied as have the impacts of fisheries on fish populations, however fewer studies have then linked fishing pressure to seabird demography. Ecologically this is fundamentally challenging to study, in part due to the time-lags in population level responses for seabird populations due their late sexual maturity, which should be accounted for in analyses studying drivers of seabird abundance (Sandvik et al., 2012). Meta-analysis type studies, particularly when modelling multiple taxa have often not accounted for this, which could lead to spurious results; this could be an explanation for one of the studies considered in our analysis that found limited evidence for fishery impacts on seabird populations (Free et al., 2021).
Most of the studies included in our analysis considered either productivity or population size, which reflects these being the most easily monitored population traits in seabird populations. However, to gain a more detailed understanding of population drivers there is a need to consider further demographic factors. While productivity and adult survival are often considered as independent traits, there is increasing evidence for carry-over effects between breeding and subsequent over-winter survival leading to these traits being correlated. For example, under low prey abundance during breeding Black-legged Kittiwake had lower over-winter survival (Kitaysky et al., 2010). Similarly in Common Guillemot, in years of high spring sea temperature (which is negatively correlated with various measures of prey quality), fledging success of chicks was reduced and subsequent overwinter survival for the adults birds (Wanless et al., 2023).
The extent to which prey availability, which is a key assumption behind reduced fishery pressure benefitting seabirds, affects seabird populations is context specific. In populations where top-down population processes dominate (e.g. predation) there may be limited, if any, impact from reduced fishing pressure. For example, Pettex et al. (2015) studied how prey availability affected colony establishment and growth in Gannet in Norway, they found that prey did not appear to be limiting factor, with disturbance and/or predation by White-tailed Eagle being the key driver of colony changes.
5.2.5.1 Knowledge gaps and future directions
The relationship between fish abundance and seabird population demography is complex and will rarely follow a linear relationship. Significant negative impacts on seabird populations generally only occur below a lower critical threshold (Cury et al., 2011; Smith et al., 2011; Guillemette et al., 2018). Additionally, seabirds are not solely impacted by prey abundance, but also the quality (e.g. lipid content) (Wanless et al., 2005a; Österblom et al., 2006; Österblom et al., 2008) and the availability of prey (Scott et al., 2006). Spatially explicit ecological models parameterized by empirical data can allow such relationships to be explored to better understand how changes in fisheries management targets could impact seabird populations (Hentati-Sundberg et al., 2021b).
The seas around Scotland are experiencing some of the most rapid rates of warming globally, with the North Sea warming by 1.3 ℃ between 1982-2006 (Belkin, 2009). This warming is associated with changes in the zooplankton community which is leading to reduced energy availability for forage fish (Olin et al., 2022). These ecosystem changes could mean that changes in fishery management may have less benefit to seabirds, if bottom-up processes become the predominant drivers of prey availability, however, conversely it has also been demonstrated that fisheries can interact with climate change to reduce fish population resilience increasing the chance of abrupt ecosystem change (Möllmann and Diekmann, 2012). Better understanding how climate change affects marine ecosystems around Scotland would improve our ability to predict how changes in fishery management could affect prey fish, and thus seabird populations, over the longer-term.
5.2.6 Conclusion
Overall, we found that prey fisheries management can benefit seabird populations. However, prey availability to seabirds is determined by many different factors (including climate related factors), with fisheries one amongst these. Whether a population level benefit can be anticipated following changes in fisheries management for a seabird species will require careful consideration of the specific ecological context, and it will rarely be possible to confidently form quantitative predictions on the level of benefit (if any). Current fisheries management rarely fully considers ecosystem impacts, including on seabird populations (ICES, 2023a), therefore, there is potential for seabird populations to benefit from changes to management approaches. Depending on how such management changes are implemented these may have potential in the context of strategic compensation.
5.3 Ecological feasibility: End of the Gannet harvest at Sula Sgeir
5.3.1 Summary
The systematic literature review and respective analyses provide evidence that, with a high degree of confidence, the cessation of chick and/or egg harvest has a low to medium effect on at least one demographic parameter of seabirds. We consider this conservation action to be less suitable as strategic compensation, as it would primarily benefit individuals from specific SPAs (i.e. Sula Sgeir in this case), resulting in potentially low overall impact at the Scottish and UK level.
5.3.2 Introduction and background
Historically, seabirds have been harvested worldwide for their eggs, meat, and down as a means of subsistence, recreation, and commercial gain (Merkel and Barry, 2008; Naves and Rothe, 2023). For certain cultures, this activity continues to represent a crucial element of their cultural heritage (Baldwin, 2012; Fyfe and Davis, 2015; Jones et al., 2015; Henri et al., 2020). However, harvests have been identified as a threat to some seabird populations (Le Corre and Bemanaja, 2009; Chen et al., 2015; Mondreti et al., 2018) and have played an important role, alongside other factors (e.g. environmental conditions, food supply, predation), in the decline, extirpation, and extinction of others, such as Little Auks Alle alle (Jakubas et al., 2022), Great Auks Pinguinus impennis (Serjeantson, 2001; Thomas et al., 2019), and Chinese Crested Terns Thalasseus bernsteini (Chen et al., 2015). For example, population viability analyses on the now-extinct Great Auk revealed that even if it had not been under threat by environmental changes, human hunting alone could have been sufficient to cause its extinction (Thomas et al., 2019).
Seabirds are one of the most threatened group of birds and many populations are rapidly declining (Croxall et al., 2012). They are highly sensitive to human exploitation as they have slow population growth; they are mostly long-lived, have high adult survival rates but low immature survival, have delayed maturity and can, when conditions are poor, skip breeding seasons (i.e. sabbaticals), and they produce small clutch sizes (Schreiber and Burger, 2001; Young and VanderWerf, 2023). This life history strategy makes seabird populations particularly susceptible to exploitation of adults. Species like Gannets, that nest colonially, have high philopatry, breed synchronously and on the ground where eggs, and chicks can be accessed easily, are particularly vulnerable to human harvest (Rodríguez et al., 2019).
Although seabird harvesting is not as important for sustenance as it was in the past, it still has an important traditional and cultural importance (Naves, 2018). Overharvesting can lead to declines in seabird populations, which can have cascading effects on the marine ecosystem. Therefore, egg and chick harvest are strictly regulated by international and national laws and is only permitted in a few places around the world, including Scotland (Denlinger and Wohl, 2001; Merkel and Barry, 2008; Naves and Rothe, 2023). The harvest is usually done by collecting eggs or chicks from nests during the breeding season, but this varies depending on the species and location.
Harvest regulations aim to manage harvests at sustainable levels by balancing harvest quantities and population size, accounting for other factors that may be affecting the populations (e.g. density-dependent processes, predation, bycatch, food availability) (Naves and Rothe, 2023). Therefore, the degree of the harvest (the number, proportions and type of harvest target i.e. eggs, chicks or adults), and the timing of the harvest within the year and within the life cycle of the species, are main factors in determining the potential impacts that harvests have on the targeted and surrounding populations (Hunter and Caswell, 2005; Moller et al., 2009; Lyver et al., 2015; Naves and Rothe, 2023).
The practice of harvesting seabirds in Scotland, primarily for their meat, eggs, oil and fat, dates back to prehistoric times (Best and Mulville, 2016). Even in 1860, seabirds and their eggs continued to play an important role in the economy and culture in several localities throughout the country (Baldwnn, 1974; Baldwin, 2012). In the 19th century, harvesting became more regulated. Complete protection to seabirds was granted in Britain by the Protection of Birds Act, introduced in 1954 but the Wild Birds (Gannets on Sula Sgeir) Order 1955 allowed members from the community of Ness, on the Isle of Lewis, to continue the Gannet harvest (Benn et al., 1989).
Sula Sgeir is a small, uninhabited rocky islet located 18 km west of North Rona (59°5’43.44" N, - 6°9’ 22.6188"W), with a large Gannet breeding colony (Angus and Maclennan, 2015). To date, 2000 Gannet chicks or ‘gugas’, equivalent to at least 17% of annual chick production, can be harvested annually – a quota set by the Scottish Government, with advice from NatureScot (formerly known as the Scottish Natural Heritage) (Wanless et al., 2015). The annual harvest occurs in late August and early September, after most seabird species have ceased breeding reducing potential for disturbance of other species, however, Fulmars (Benn et al., 1989; Angus and Maclennan, 2015) and Storm Petrel will still be present. The harvest itself is believed to be sustainable (in terms of not leading to a population decline), as the 2013 count indicated that the number of Apparently Occupied Sites increased over the previous nine years despite the continued harvesting. This slight increase may, however, be due to immigration of individuals originating from St Kilda and/or Sule Stack (Wanless et al., 2015), and these studies were prior to recent impacts (2022 and 2023) from HPAI which heavily affected Gannet populations across the UK (Tremlett et al., 2024).
Currently, the UK holds 56% of the world’s breeding Gannet populations, the majority of which nest on cliffs or islands in Scotland (JNCC, 2021). It is important to recognise the cultural importance of the harvest, while also working to minimise the negative impacts this may have on Gannet populations. This literature review aims to gather evidence on the impact that egg and chick harvests have on seabird populations to gain a clearer understanding of the possible impacts that the 2000 chick quota may be having on the Sula Sgeir and surrounding Gannet populations.
5.3.3 Methods
The corresponding research question to this conservation action is: ‘Would the Gannet population at Sula Sgeir and/or in the wider SPA network benefit from ending the harvest of Gannet chicks at Sula Sgeir?’
A detailed description of the research question, keywords, search strings, and study selection (Figure 13) is provided in section 3 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 13 references were included for the final review.
5.3.4 Results
5.3.4.1 General observations
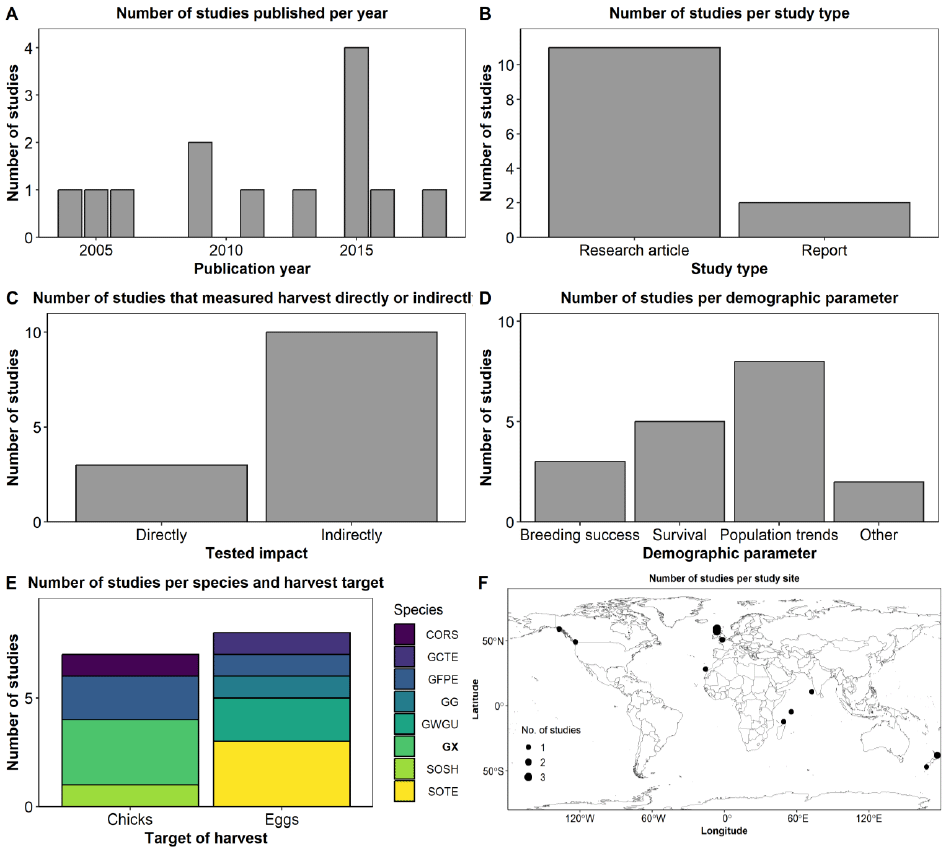
We identified 13 references that met our criteria of measuring the effect of chick and/or egg harvest on seabird populations. All of which were published in English. The literature review yielded publications ranging from 2004 to 2018 (Figure 14A). The median publication date is 2013. The year 2015 had the highest number of publications among the identified references. References were either research articles or reports, representing 85% and 15%, respectively (Figure 14B). No review papers were included for the final assessment. Most studies (77%) measured or assessed the impact of chick and/or egg harvest indirectly, either by employing population models or by simulating population trends under different levels of harvest intensity rates (Figure 14C). On the other hand, 22% of the studies used empirical data to compare demographic parameters between harvested and neighbouring unharvested colonies with seemingly comparable environmental conditions (Figure 14C).
Most studies (62%) analysed or modelled the impact of chick and/or egg harvest on population trends, 39% on survival rates, 23% on productivity or breeding success, and 15% on other parameters such as predation rates and egg quality (Figure 14D). Half of the studies explored the effect of chick harvest and the other half the effect of egg harvest. Note that Figure 14E does not reflect the latter statement, as one reference studied the effect of egg harvest on two different seabird species. Studies focussed on several seabird species (listed from most studied to least): Sooty Tern (Onychoprion fuscatus; ‘SOTE’), Gannet (‘GX’), Grey-faced Petrel (Pterodroma gouldi; ‘GFPE’), Glaucous-winged Gull (Larus glaucescens; ‘GWGU’), Cory’s Shearwater (Calonectris borealis; ‘CORS’), Great Crested Tern (Thalasseus bergii; ‘GCTE’), Sooty Shearwater (Ardenna grisea; ‘SOSH’), and Black-headed Gull (Chroicocephalus ridibundus; ‘GG’) (Figure 14E). All Gannet-related studies explored the effect of chick harvest. Studies collected, analysed, or simulated data from the UK, Spain, Madagascar, Seychelles, India, New Zealand, USA, and Canada (Figure 14F). In the UK, three records investigated the population trend of the Sula Sgeir Gannet population (Furness et al., 2013; Murray et al., 2015; Trinder, 2016), and one study explored the impact of egg harvesting on breeding success of Black-headed Gulls in Hampshire and Dorset in southern England (Wood et al., 2009).
The most frequent data collection methods were observational field data, nest monitoring, aerial and terrestrial surveys, harvest observations, ringing, questionnaires, and interviews. Numerous studies utilised population models and simulations as analytical tools. Most of the authors contributed to only one of the included references.
5.3.4.2 Evidence scoring
The relevance, as well as the ecological efficacy scores for all 13 references are shown in Table 24 and Figure 15 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Overall, references were of medium relevancy for our assessment, with only three studies surpassing 20 points. Scores ranged between 13 and 26 out of a maximum possible score of 30, with a median and mean score of 17 and 18, respectively. For this reason, studies varied in their associated weight, ranging from 0.057 to 0.114 (Table 24).
The ecological efficacy, i.e. the likelihood that ceasing chick and/or egg harvest is beneficial on at least one seabird demographic parameter, varied between studies. Out of a maximum possible score of 25, scores ranged between 3 and 16. The median score is nine. This conservation action, however, is peculiar. Any degree of chick or egg harvest will undoubtedly have detrimental effects on seabird populations. This is because individuals are being taken out artificially and not replaced. The only exception to this is when eggs are harvested at the start of the season, provided that the birds relay. Therefore, it is important to note that the focus of most studies was not on testing the direct effect of the conservation action itself, ending the harvest, but rather on measuring or simulating the impact of the effect of the harvest on seabird populations. We, therefore, assumed that a strong negative effect of harvest would indicate a high beneficial effect if the conservation action were to be implemented. Hence, to estimate the degree of the effect we considered the proportion of chicks and/or eggs being harvested, the degree of the effect discussed by authors, and the status of the species. The low to medium scores reflect the degree of statistical inference (e.g. weak statistical power) rather than the degree of effect.
Table 24. Relevance and ecological efficacy scores of each assessed study on ending chick and/or egg harvest review. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of ending the chick and egg harvest, or those with low statistical inference; higher scores reflect studies that found a strong beneficial effect of ending harvests with high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
The sum of all final weighted scores is 9.4 (i.e. the mean of ecological efficacy weighted by relevance), therefore, it can be concluded that the evidence overall support that ending chick and/or egg harvest will benefit seabirds to a low-medium degree (Figure 15), although, as discussed previously, the proportion of chicks and/or eggs being harvested and the conservation status of the affected species will play a major role in this effect.
5.3.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this conservation action was 11 out of the possible 15 (Table 25), suggesting that we have a high degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of ending harvests.
Table 25. Assessment of the confidence in the evidence for ending chick and/or egg harvests.
This table can be viewed in Excel format.
5.3.4.4 Main findings: the effect of ending Gannet chick and/or egg harvest
In summary, our literature review and respective analyses provide evidence that, with a high degree of confidence, the cessation of chick and/or egg harvest has a low-medium effect on at least one demographic parameter of seabirds, but this is highly dependent on the proportion of the population being harvested, the conservation status of the species, and the timing of the harvest within the year and within the life cycle of the species.
The key findings from the most relevant studies are summarised in Table 26. Overall, chick and/or egg harvest is shown to negatively impact seabird populations. For example, one study investigated the changes in Glaucous-winged Gull populations over the course of a century (Blight et al., 2015) and suggest that the strong population increases observed during the mid-1900s correspond with the passing of the 1917 Canadian Migratory Birds Convention Act that led to the cessation of seabird harvests. Similarly, in India, fishermen were estimated to be harvesting 14–45% of the eggs of an already declining species, the Sooty Tern. At this rate, this unregulated and illegal activity may drive this population to extirpation in the near future (Mondreti et al., 2018).
Studies that directly tested differences between harvested and unharvested sites also found significant differences in breeding success, population trends, and overall survival rates between colonies. In Madagascar, tern colonies were monitored to understand the impact of human harvest. For a colony of Sooty Terns that was intensively harvested for eggs; it was estimated that 66% of the eggs were harvested. Although regulated, the harvest rate is probably beyond the sustainable threshold and may lead to declines (Le Corre and Bemanaja, 2009). On the other hand, a colony of Great Crested Terns increased 10-fold, from 1000 pairs in 1997 to 10840 in 2008 when it became protected, and harvests stopped completely (Le Corre and Bemanaja, 2009). Similarly, on the Canary Islands, the breeding success of Cory’s Shearwaters was higher in unharvested colonies compared to harvested colonies, and the colonies with low to medium harvest intensities had higher breeding success than those intensively harvested (up to 63% of chicks) (López-Darias et al., 2011). In southern England, those sites where Black-headed Gull eggs were harvested were characterised by reductions in egg volume, yolk-to-albumen ratio, and eggshell thickness, which translated to poorer breeding success and chick survival (Wood et al., 2009).
Table 26. Summary findings of the five most relevant studies on the effect of ending chick and/or egg harvests, in order of decreasing relevance.
This table can be viewed in Excel format.
Although harvests have proven to have a detrimental effect on populations, many studies highlight the cultural and social importance of seabird harvesting for certain cultures around the world. These studies argue that sustainable harvesting practices may even allow for sustained population growth rates and are not necessarily synonymous with declines. For example, Grey-faced Petrels in New Zealand, could potentially sustain a fixed-quota harvest of up to 6,000 chicks or a fixed-proportion harvest of up to 30% of chicks before the population is predicted to decline (Lyver et al., 2015). Harvesting at a fixed intensity but less frequently, earlier in the season and at a fixed quota could further reduce the degree of effect (Feare and Doherty, 2004; Zador et al., 2006; Jones et al., 2015).
Regarding the Gannet population at Sula Sgeir, Trinder (2016) suggested that the harvest has reduced the rate of population growth below the level that would be predicted in the absence of a harvest, and this has likely impacted other neighbouring populations linked through immigration and emigration, such as those from St Kilda and Sule Stack. Nevertheless, the Sula Sgeir population is increasing, and there appears to be space for the colony to expand further, suggesting that the population may not be at maximum carrying capacity with respect to nest sites. In fact, the number of Apparently Occupied Nests increased 22% from 2004 to 2013 (Murray et al., 2015; Wanless et al., 2015). This steady increase could continue despite the current harvest level (of 2,000 chicks per year), but levels above 3,500 chicks would likely lead to long-term declines (Trinder, 2016). Ending the chick harvest would increase productivity and would likely result in more rapid growth of breeding numbers. Furness et al. (2013) concluded that the termination of the harvest would be the most cost-effective approach to increasing survival or productivity for Gannet at the level of the British Isles.
5.3.4.5 Biases or Conflict of interest
The authors did not disclose any biases or conflicts of interest, and our investigation did not reveal any such concerns.
5.3.5 Discussion
All the literature reviewed on this subject agreed that harvesting chicks and eggs undoubtedly affects seabird populations. The degree of this effect, however, depends significantly on the harvest target, the proportion of the population being harvested, the conservation status of the population, the timing of the harvest within the year and within the life cycle of the species, and other population-limiting factors, such as environmental conditions, predation rates, pollution, and diseases.
Gannet survival rates increase with age. In Britain and Ireland, for example, first-year Gannets have an annual survival rate between 0.41 and 0.44, second-, third-, and fourth-year birds have an annual survival rate between 0.82 and 0.90, and adult birds have an annual survival rate of 0.92 (Wanless et al., 2006). The differential survival rates between young and adults, mean that the target of the harvest, whether it is eggs, chicks, or adults have varying effects on population dynamics. Egg harvests, if timed correctly and undertaken earlier in the season, may be less disruptive than chick and adult harvest, as birds can relay and still produce fledglings, depending on the timing of harvest and on food availability (Wood et al., 2009). In contrast, when chicks are harvested, relaying is not possible (it being too late in the season), and birds cannot compensate for the extraction.
The population growth rate of Gannet, like many other long-lived species, is much more susceptible to changes in adult survival than reproduction (Hunter and Caswell, 2005), therefore, adult harvests result in significantly worse population level consequences than both egg and chick harvests. Models using data gathered from Grey-faced Petrels estimated that the population started to decline when 2% or more of adult birds were harvested compared with the threshold harvest of 25% or more of either eggs or chicks (Lyver et al., 2015). Similarly, in Sooty Shearwater a chick exploitation rate of 5% reduces population growth by 0.3%, whereas a 5% adult exploitation reduces population growth by 3% (Hunter and Caswell, 2005). The fact that adults are not harvested in Madagascar may explain the persistence of a large Sooty Tern colony, despite a significant egg harvest of up to 66% (Le Corre and Bemanaja, 2009).
How harvests are managed – the proportion of the population being harvested and the timing and intensity of the harvest within the year – will also determine the degree of effect of the harvest on bird populations. Several studies have identified population- and species-specific sustainable harvest thresholds; 5% of chicks for Sooty Shearwaters (Hunter and Caswell, 2005), 20% of eggs for Glaucous-winged Gulls (Zador et al., 2006) and Sooty Terns (Feare and Doherty, 2004), and 6000 chicks, or 30% of chicks or eggs, for Grey-faced Petrels (Jones et al., 2015; Lyver et al., 2015). Similarly, harvesting at a fixed intensity, with less frequency, and earlier in the season may reduce the negative impacts on reproductive success by increasing the probability of relaying, reducing disturbance, other types of predation, and nest abandonment, and increasing breeding synchrony (Zador et al., 2006; Lyver et al., 2015). However, species of higher conservation concern, especially those with declining populations, should not be harvested under any circumstances, as even relatively small harvests can have detrimental effects on these populations (Chen et al., 2015; Alfaro-Shigueto et al., 2016).
5.3.5.1 Gannets at Sula Sgeir
Gannet lay a single-egg clutch and do not replace lost clutches, therefore, if management regimes were similar, egg and chick extraction may potentially have the same effect on the Sula Sgeir population (except in terms of carry-over effects, see e.g. (Fayet et al., 2016)). Although Trinder (2016) predicted that the current harvest regime has resulted in a small reduction in the population growth rate compared to what would be expected without the harvest, the population had been increasing (2.2% in the last decade) up to that assessment, and Trinder concluded that this growth was likely to continue with the harvest level at that time. As long as the harvest threshold is below 3500 chicks per year, immigration is stable, and environmental conditions remain relatively similar, then the Sula Sgeir population was expected to remain stable or to increase (Trinder, 2016). Given the above, and that Gannet population growth rates are more sensitive to changes in adult survival rates, rather that immature survival rates and breeding success (WWT Consulting, 2012), the Sula Sgeir chick Gannet harvest seems to have had a relatively small effects on Gannet population size over past years.
However, Trinder (2016) proposed that the level of growth observed at Sula Sgeir is only possible in the face of current harvest levels with immigration of Gannet from other colonies. It was calculated that the population requires over 270 breeding age recruits annually from neighbouring colonies, such as St Kilda and Sule Stack. This inevitably means that the harvest reduces the number of recruits available for other colonies, especially those located nearer to Sula Sgeir, although, as Scottish Gannet populations increase, an overall increase in potential recruits is expected. Additionally, at a meta-population level, chick and egg harvesting can lead to population fragmentation and loss of genetic diversity (Naves and Rothe, 2023). As certain colonies are targeted for harvesting, the genetic makeup of the population may become increasingly homogenous, making it more vulnerable to environmental stressors and reducing the overall resilience of the species.
It is unclear how the harvest affects other UK colonies, but the impact, if any, should diminish as the distance from the harvesting site increases (see e.g. Jeglinski et al. (2023)). Ending the chick harvest at Sula Sgeir would enhance breeding success at the colony and potentially accelerate the growth of neighbouring Gannet populations, but may not necessarily lead to significant changes in colonies farther away (e.g. North East Scotland, England).
It is important to note that the impact of harvests is influenced by other factors that limit population growth, including predation rates, pollution, disease, bycatch, and environmental conditions at breeding and non-breeding sites. Therefore, if the harvest were to continue, then the harvest regime, which involves a 2000 chicks per year quota, should be evaluated periodically, and carefully adjusted based on current population trends and environmental factors. This is particularly important given that stochastic events, such as the 2022 outbreak of Highly Pathogenic Avian Influenza (HPAI), can have significant detrimental effects on Gannet (Tremlett et al., 2024; Jeglinski et al., preprint). If other limiting factors are causing population growth to decline, ending, or reducing, the harvest should be considered.
5.3.5.2 Knowledge gaps and future directions
Assessing the relative impacts of harvest and other threats on seabird populations is a challenging task that requires a substantial amount of data and appropriate population modelling frameworks, which are often scarce. Although Before-After-Control-Impact (BACI) studies would be the preferred approach for understanding the impact of ending seabird harvests, they are not always feasible due to illegal harvesting and the difficulty of accurately estimating harvest rates. Additionally, we could not find any records that have measured the effect of ending the harvest in a single colony. It is important to note that population models and simulations are typically used to predict the impact of harvest on seabirds, but they have limitations; the accuracy of these models are highly dependent on the data available, and the parameters used, which are often incomplete, and the degree of uncertainty increases over time when testing the models with multiple years of data. Furthermore, these models rarely model meta-population processes (e.g. immigration/emigration and density dependent processes), although this could now be explored further for Sula Sgeir by building on a recently developed meta-population model by Jeglinski et al. (2023) and Peery et al. (2006). This would be particularly valuable to conduct now that we have updated population counts for key Gannet colonies (including Sula Sgeir) following the HPAI outbreak in 2022/2023 (Tremlett et al., 2024).
It is crucial to recognise that the survival of seabird populations, such as the Sula Sgeir population, is not solely impacted by harvest, but by other factors as well, including bycatch during the breeding period and migration, and HPAI. To make informed decisions about harvest quotas, it is essential to assess the impact of these factors on the population and neighbouring populations and adjust the harvest quota accordingly.
5.3.6 Conclusion
This conservation action is less suitable as strategic compensation since it would provide primarily local benefits (i.e. Sula Sgeir, Sule Stack), limited at the regional level due to metapopulation dynamics, and would likely have a low impact at Scotland and UK level. If the harvest were to continue, regular reviews of the level of harvest should take place, accounting for current stochastic events, such as the HPAI, and involving collaboration between researchers, government, and harvesters.
Any type of seabird harvest will undoubtedly affect seabird populations. The degree of this effect, however, depends significantly on the harvest target, the proportion of the population being harvested, the conservation status of the species, the timing of the harvest within the year and within the life cycle of the species, and other population-limiting factors, such as environmental conditions, predation rates, pollution, and diseases.
The Sula Sgeir Gannet population (the impact of the harvest at Sula Sgeir is considered further in the relevant section in WP2) is subject to a licensed annual harvest of 2,000 full-grown chicks. The harvest occurs over a two-week period towards the end of the breeding season and is restricted to accessible areas. At this stage, most chicks are already well-developed, so are less vulnerable to disturbance meaning that indirect mortality should be low (i.e. mortality to additional chicks beyond those harvested), though levels of additional mortality are not known. Given that the Sula Sgeir Gannet population, as well as most other Scottish Gannet populations, has been increasing[3] and adult mortality has a stronger effect on population growth than breeding success, the harvest does not appear to have had a strong impact on the population persistence but may be affecting the overall population size. While discontinuing the controlled harvest would be expected to accelerate the growth rate of the Sula Sgeir colony and to a lesser extent that of neighbouring populations, the impact at the UK scale is likely be minimal due to the distance and small levels of emigration to Sula Sgeir. In addition to ecological considerations, there are also cultural heritage considerations to ending the harvest, given that the traditional harvest has persisted for centuries.
5.4 Ecological feasibility: Mammalian predator eradication and/or management
5.4.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, the eradication and/or control of invasive mammalian predators has a medium effect on at least one demographic parameter of seabirds. The degree of success, however, will depend on the focal seabird species, the mammalian predator that is being eradicated/controlled, the success of post-management biosecurity measures, and external factors (e.g. distance to mainland and other islands, climate change, presence of other predator species, terrain, and level of human activity).
5.4.2 Introduction and background
Invasive alien species (also called invasive non-native species), as defined by the Convention on Biological Diversity (CBD, 2023), are species introduced by humans, intentionally or unintentionally, outside their natural geographic range. These species have a significant impact on native biological diversity in the areas they invade. They share common characteristics such as rapid reproduction and growth, high dispersal ability, generalist foraging strategies, and the capability to thrive in diverse and novel environments. Unlike in their native habitats, invasive species often encounter a lack of natural predators and competitors in the introduced areas, allowing them to flourish and reproduce with minimal constraints (CBD, 2023).
Invasive species are the primary land-based threat to seabird species worldwide (Phillips, 2010; Dias et al., 2019; Spatz et al., 2023a). Seabirds are primarily impacted by invasive species through direct predation at the breeding sites (usually of eggs and young), especially on islands, but are also impacted indirectly through habitat degradation, stress, trampling of nests, disease transmission, and competition. Seabirds have life histories characterised by longevity and low fecundity which make them slow to recover from predation and chronic reproductive failures caused by invasive species. Seabirds have not evolved to respond to terrestrial predation by vertebrate predators, and many seabirds nest in colonies on the ground or in burrows and crevices rendering them more susceptible to predation. In particular, predation by invasive mammals has resulted in the probable global extinction and local extirpations of several seabird species and has contributed to the globally threatened status of many other species (Blackburn et al., 2004).
Predation by invasive mammals, such as rats and cats, is the primary driver behind the decline of seabird populations worldwide (Dias et al., 2019). These species have successfully established themselves on most major islands around the world. Cats and rats, in particular, pose a significant threat due to their ability to target all life stages of seabirds and their proficiency in accessing and preying upon seabird nests, irrespective of their remoteness or inaccessibility (Angel and Cooper, 2006; Jones et al., 2008; Le Corre, 2008). Nonetheless, other medium-sized mammals such as mink, dogs, goats, and foxes have also caused detrimental effects on seabirds (Moore et al., 2003; Davis et al., 2018). The degree of vulnerability to invasive mammals depends on the seabird species according to its size and nesting ecology (Lewison et al., 2012). For example, while nearly all seabird species can be preyed upon by rats, the impact is stronger among smaller species that nest in crevices or burrows compared to those nesting on cliff-tops (Jones et al., 2008).
The eradication and/or control of invasive mammalian species have proven to be an effective conservation strategy for seabird conservation (Genovesi, 2005; DIISE, 2018). There is increasing evidence that implementing eradication and/or control measures targeting invasive mammals can yield substantial benefits for seabird populations breeding on islands. Eradication involves the complete removal of wild populations of invasive species from a defined area, presenting an opportunity for significant long-term benefits. On the other hand, control measures aim to suppress the invasive species population through ongoing removal and control efforts. The decision to pursue eradication or control measures depends on the specific predator and the site where management is being considered. To date, more than 2000 efforts to eradicate invasive vertebrate species, mostly mammals, have taken place on over 800 islands around the globe in 60 countries, with an overall success rate of 88% (DIISE, 2018). These efforts have had significant benefits to seabirds, from increases in productivity and survival, to population growth, distribution expansion, and recolonisation. Jones et al. (2016) reported that 84 out of 87 seabird species exhibited a beneficial trend following mammal eradication on islands.
Scotland, with its abundance of islands, is not exempt from the challenges posed by invasive mammal species. The presence of such species (often multiple species) is confirmed or suspected for many of Scotland’s islands (Stanbury et al., 2017). The introduction of rats, cats, mink, and other invasive mammals (invasive here means being present outside of their normal range, e.g. many islands are historically free of rodents) to these islands has had severe detrimental effects on local seabird populations. The aim of this literature review is, therefore, to identify the main mammalian predators in Scotland and gather all available evidence of the effect of mammalian predator eradication and/or control efforts on seabird species, with a focus on our focal species. By examining the effectiveness of predator eradication and/or control efforts, conservation efforts can be better targeted and prioritised to find the most cost-effective solutions, which may have potential as strategic compensatory measures of the impacts of offshore wind farms on seabirds. Additionally, this review aims to identify which seabird species would benefit most from predator eradication or control efforts to strategically target eradication efforts to help protect and conserve our focal seabird species (i.e. those species most likely to be affected by offshore windfarms).
5.4.3 Methods
This conservation action was associated with two research questions. The first, ‘What is the potential for seabirds to have increased productivity or survival from mammalian predator eradication/management?’ explores the effect of eradication and/or control of mammalian predators on seabird populations. Then the second question, ‘Among mammalian predators, which ones offer the most potential for effective eradication and/or management?’, aims to identify the mammalian species with higher probabilities of eradication and/or management success.
A detailed description of the research question, keywords, search strings, and study selection (Figure 16) is provided in section 5 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
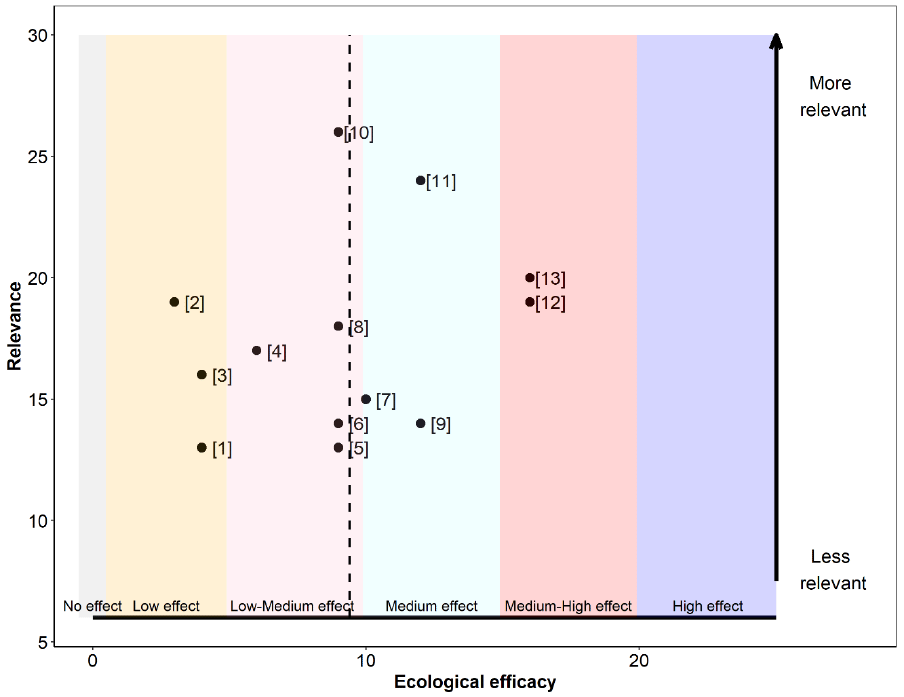
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 23 references were included for the final review.
5.4.4 Results
5.4.4.1 General observations
We identified 23 references that met our criteria of measuring the effect of eradication and control of mammalian predators on seabird populations, all published in English. The literature review yielded publications ranging from 2001 to 2021 (Figure 17A). The median publication date is 2010. The years 2006, 2008, and 2019 had the highest number of publications among the identified references. Most references were research articles (78%), but we also identified two research note articles, a review, a report, and a scientific blog post (Figure 17B). Most studies (96%) measured or assessed the impact of mammal eradication and/or control directly, meaning that the impact on seabirds was measured empirically either by before-and-after studies, or by comparing managed and non-managed sites (Figure 17C).
Studies researched the effect of several mammalian predators on diverse seabird species groups (Figure 17D). Out of the studied mammalian predators, Black rats had the highest number of studies focussed on their eradication and/or control, with 12 studies. Brown rats followed with nine studies, then mink with four, and cats and foxes with two and one study, respectively. Studies focussed on several seabird species groups (listed from most studied to least): petrels and shearwaters (13 studies; i.e. Cory’s Shearwater, Manx Shearwater, Audubon’s Shearwater, Yelkouan Shearwater, Wedge-tailed Shearwaters, Black-vented Shearwater, European Storm Petrel, Band-rumped Storm Petrel), auks (8 studies; i.e. Atlantic Puffin, Razorbill, Black Guillemot, Ancient Murrelet, Cassin’s Auklet, Scripp’s Murrelet), gulls (6 studies; i.e. Black-legged Kittiwake, Great Black-backed Gull, Lesser Black-backed Gull, Herring Gull, Common Gull), terns (5 studies; Arctic Tern, Common Tern, Little Tern, Sooty Tern, White Tern), fulmars (3 studies), European shags (2 studies), and 5 studies focussed on other seabird species (i.e. Ascension Frigatebird, Magnificent Frigatebird, Brown Noddy, Black Noddy, Arctic Skua, Masked Booby, Brown Booby, Red-footed Booby, White-tailed Tropicbird, Red-billed Tropicbird). Note that some reference studied multiple predators and seabird species.
Of the studies analysed, 57% focussed on the impact of mammal eradication/control on breeding success, while 30% studied breeding pairs/population, 26% studied recolonisation, 9% studied survival/population trends, and 13% focussed on other parameters such as population size and burrow occupancy (Figure 17E). Most studies (65%) focussed on mammal eradication, with 39% studying control (one study analysed both) (Figure 17F). Studies were conducted primarily on islands (96%), with only one study on foxes conducted on the mainland. Data was collected, analysed, or simulated from multiple islands across the globe, including the UK and its overseas territories, Canada, French territories in the Indian Ocean, Finland, Malta, USA, Spain, and Mexico (Figure 17G). Within the UK, nine eradication and control projects were carried out, primarily on islands (Figure 17H). These projects included various sites on the west coast of Scotland, such as Handa Island, the Island of Rum, the Uists, Canna Island, Sanday Island, and Ailsa Craig, along with Lundy Island and South Walney in England, and Ramsey Island in Wales (Figure 17H). Of these studies, six examined the impacts of rat eradication, two focussed on mink control, and one on fox control.
The most frequent data collection methods were observational field data, nest monitoring, and surveys. The most frequently employed mammalian eradication methods were poison baiting, followed by traps and shooting. Most of the authors contributed to one reference.
5.4.4.2 Evidence scoring
The relevance, as well as the ecological efficacy scores for all 23 references are shown in Table 27 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Overall, references were of medium-high relevancy for our assessment, with ten studies surpassing 20 points. Scores ranged between 12 and 28 out of a maximum possible score of 30, with a median and mean score of 20. For this reason, studies varied in their associated weight, ranging from 0.026 to 0.061 (Table 27). Out of the assessed studies, all but one suggests that eradication/control efforts had a positive effect on at least one demographic parameter of seabirds. However, the study that did not observe any effect attributed this to the presence of other rodents on the island that began predating seabirds after the target rodent (rats) was eradicated (Lambert et al., 2021).
The ecological efficacy, i.e. the likelihood that eradication/control efforts are beneficial on at least one seabird demographic parameter, varied across studies. Scores ranged between 3 (eradication/control efforts had a low effect on seabirds) and 20 (eradication/control efforts had a strong effect on seabirds) out of a maximum possible score of 25. The median score was 12, indicating that the effect varied according to the study. Half of the studies found a low to medium effect, while the other half found a medium to high effect.
The sum of all final weighted scores is 12.8 (i.e. the mean of ecological efficacy weighted by relevance), indicating that overall, the evidence suggests that mammalian eradication and/or control will benefit seabirds to a medium degree (Figure 18), although, as we will discuss, the degree of this effect is influenced by factors such as the species, of both the predators and the seabirds, the future biosecurity measures and on external factors.
Table 27. Relevance and ecological efficacy scores, of each assessed review. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of eradicating/controlling mammalian predators, or those with low statistical inference; higher scores reflect studies that found a strong beneficial effect of eradicating/controlling mammalian predators with high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
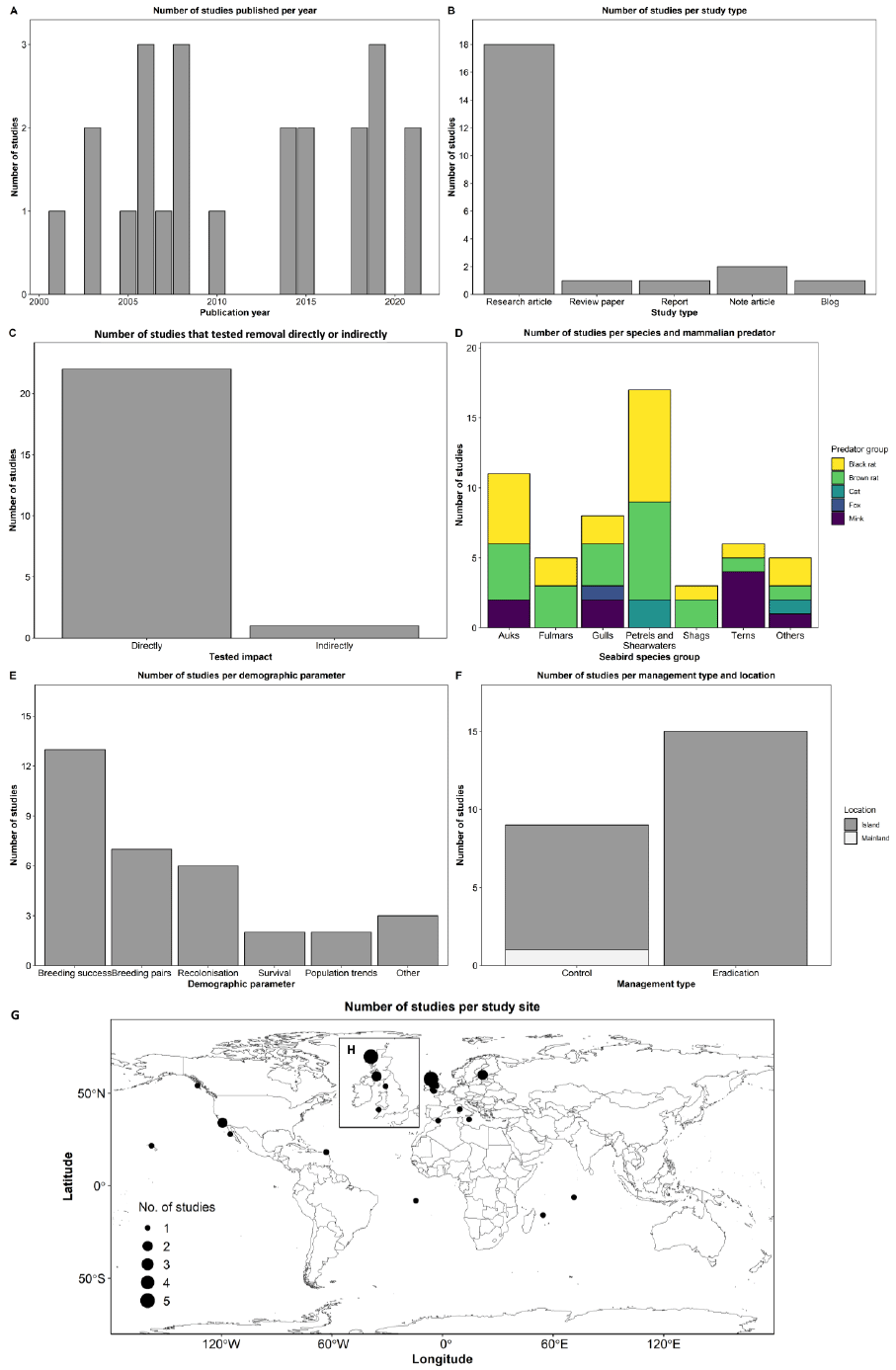
5.4.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this conservation action was 10 out of the possible 15 (Table 28), suggesting that we have a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of mammalian predator eradication/control efforts.
Table 28. Assessment of the confidence in the evidence for eradication and/or control of mammalian predators.
This table can be viewed in Excel format.
5.4.4.4 Main findings: the effect of mammalian predator eradication and/or management
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, the eradication and/or control of mammalian predators has a medium effect on at least one demographic parameter of seabirds, but this depends on the seabird species, the mammalian predator that is being eradicated/controlled, the success of post-management biosecurity measures, and external factors.
The key findings from the most relevant studies are summarised in Table 29. Overall, the effect of successful eradication/control efforts had a positive effect on at least one demographic parameter of seabirds. The studies that did not find a significant benefit or found a low benefit attributed this to external factors such as other predators present in the area or environmental conditions, and not to the predator eradication/control per se (e.g. Blight et al. (2015)).
Table 29. Summary findings of the six most relevant studies on the effect of mammalian predator eradication and/or control on seabirds, in order of decreasing relevance.
This table can be viewed in Excel format.
The effect that eradication/control had on seabirds varied strongly on the predator and seabird species. A summary of the effect that eradication and/or control efforts had on seabirds are shown in Table 30. For further details and references see Table A 3.
Table 30. Key findings of the studies for each seabird species within the mammalian predator eradication and/or control on seabirds review. Focal species are highlighted in italics. Information regarding boobies, frigatebirds, noddies, and tropicbirds were excluded as it was deemed less relevant. The effect is colour-coded as follows: red or ‘=’ = no effect, white or ‘+’ = small effect but not enough to reflect a population increase (e.g. populations stabilised or recolonisation but no evidence of breeding or decreased rate of decline), and blue or ‘++’ = effect that led to a population increase (e.g. measured increase in productivity).
This table can be viewed in Excel format.
A total of 16 studies researched the effects of Brown and/or Black Rats on seabird populations, all were conducted on islands across the globe (i.e. rather than mainland sites). These islands ranged in distance from <1 km to 3,200 km from the mainland. Overall, studies demonstrated that rats pose a significant threat to seabirds, and their successful eradication and/or control can have a positive impact on seabird populations (Table 30). Species such as terns (Stoneman and Zonfrillo, 2005), fulmars (Zonfrillo, 2001), Lesser Black-backed Gulls (Zonfrillo, 2001), European Shags, (Luxmoore et al., 2019), Ancient Murrelets (Regehr et al., 2007), Yelkouan Shearwaters (Lago et al., 2019), Wedge-tailed Shearwaters (Marie et al., 2014), Cory’s Shearwaters (Igual et al., 2006; Pascal et al., 2008), and Scripp’s Murrelet (Whitworth and Carter, 2018) increased breeding success. Puffins (Stoneman and Zonfrillo, 2005; Luxmoore et al., 2019), Ancient Murrelets (Regehr et al., 2007), frigatebirds (Bright et al., 2014), boobies (Bright et al., 2014), Manx Shearwaters (Bell et al., 2019), and Scripp’s Murrelets (Whitworth and Carter, 2018) increased number of breeding pairs. Kittiwake (Luxmoore et al., 2019), Common Gulls (Luxmoore et al., 2019), Manx Shearwaters (Appleton et al., 2006), Scripp’s Murrelet (Whitworth and Carter, 2018), and boobies (Le Corre et al., 2015) populations are increasing or predicted to increase. Cassin’s Auklets (Regehr et al., 2007; Whitworth et al., 2015), Storm petrels (Stoneman and Zonfrillo, 2005; Bell et al., 2019), Manx Shearwaters (Zonfrillo, 2001; Luxmoore et al., 2019), and Black Guillemots (Zonfrillo, 2001) recolonised and/or presented evidence of breeding. Herring Gulls (Luxmoore et al., 2019), Great Black-backed Gulls (Luxmoore et al., 2019), Lesser Black-backed Gulls (Luxmoore et al., 2019) populations stabilised but prior to eradication were declining. Populations of Guillemots (Luxmoore et al., 2019) and Razorbills (Luxmoore et al., 2019) are continuing to decline but at slower rates. Great Black-backed Gull (Zonfrillo, 2001), Manx Shearwaters (Lambert et al., 2021), fulmars (Stoneman and Zonfrillo, 2005; Luxmoore et al., 2019) did not show evidence of beneficial effects.
Four studies researched the effects of Mink on seabird populations, all conducted on islands within Scotland and Finland. The distance from the islands to the mainland varied but were generally close (<150 km). Due to the proximity of the islands to other islands or the mainland, and that mink can swim long distances, mink are hard to eradicate and are therefore usually controlled rather than eradicated. Results from the studies showed varying effects of mink control on seabirds. In the Scottish west coast, terns showed higher breeding success at controlled sites, compared to uncontrolled sites (Ratcliffe et al., 2006; Ratcliffe et al., 2008). In the Archipelago National Park, Finland, Mink have been controlled since the 1990s. The effect it has had on biodiversity has been substantial, with many seabirds, land birds, and water birds having benefited from it (Nordstrom et al., 2003; Banks et al., 2008). Regarding seabirds, Great Black-backed Gulls have not shown signs of change since the control, Razorbills recolonised the sites, and Arctic Skuas, Arctic Terns and Common Gulls increased their breeding success.
The two studies that focussed on feral cat eradication occurred in the tropics. In Ascension Island, a UK overseas territory, feral cats were eradicated (Ratcliffe et al., 2010). The results were highly seabird species-specific. Species that showed population increases were tropicbirds, Masked and Brown-footed Boobies, and Brown Noddies. Other species such as the Madeiran storm petrel, frigatebirds, Red-footed Boobies, and White Terns did not show evidence of population increase or recolonisation. In Natividad Island, Mexico, mortality rates of Black-vented Shearwaters decreased 90% after cat eradication (Keitt and Tershy, 2003).
The single study that focussed on Red Foxes was undertaken in South Walney, England (Davis et al., 2018). Results showed that, for both Lesser Black-backed Gulls and Herring Gulls, the predator-exclusion fence appeared to have some beneficial effect, with productivity being significantly higher than expected within the fenced areas compared to the unfenced areas. This appeared to be largely due to higher survival at the chick stage.
The impact of eradication and/or control measures varied depending on the nesting ecology of birds, as shown in Table 30. Of burrowing nesters, 88% experienced significant positive effects, one record showed a weak beneficial effect, and one record showed no effect. Regarding ground nesting birds, 83% of the records indicated a strong beneficial effect, while 11% showed a weak beneficial effect, and 6% showed no effect. Among crevice nesters, 64% experienced a strong beneficial effect, 27% a weak beneficial effect, and one record showed no effect. As for cliff nesters, 50% of the records displayed a strong beneficial effect, 30% showed a weak effect, and 20% showed no effect.
Furthermore, a review conducted by Brooke et al. (2018) examined the impact of mammal eradication efforts on seabird populations on islands. The authors collected data from the Database of Island Invasive Species Eradications (DIISE, 2018), comprising information on 181 populations of 69 seabird species across 61 islands. The results of this study demonstrate that, following successful eradication, the median population growth rate was 1.119; the number of populations exhibiting positive growth (151 populations) far exceeded those experiencing declines (23 populations), while seven populations showed no significant population change. Authors also identified that population growth was faster at: newly established colonies compared to those already established, during the first few years after eradication, among gulls and terns compared to other seabird groups, and when several invasive mammals were eradicated simultaneously.
5.4.4.5 Biases or Conflict of interest
The authors did not disclose any biases or conflicts of interest, and our investigation did not reveal any such concerns.
5.4.5 Discussion
Overall, evidence suggests that the eradication/control of invasive mammalian species has a beneficial effect on at least one demographic parameter of seabird populations at the breeding sites. These results are also supported by global assessments in recent literature reviews such as those elaborated by Croxall et al. (2012), Jones et al. (2016), Brooke et al. (2018), Sutherland et al. (2021), and Spatz et al. (2023a). These findings are also in common with previous recommendations developed around eradication and biosecurity for UK islands (Stanbury et al., 2017). The degree of this effect, however, is highly dependent on the mammalian predator, the nesting ecology of the seabird species, the success of post-management biosecurity measures, and other external factors.
5.4.5.1 Predators
Findings from this literature review indicate that the impact of eradication/control efforts varied between rats, cats, mink, and foxes, the predominant invasive mammals in Scotland, and their management approach (i.e. eradication or control).
Black and Brown rats are the invasive species with the most devastating impacts on seabird populations and are likely responsible for the greatest number of extinctions and ecosystem changes on islands caused by any predator (Towns et al., 2006; Dias et al., 2019; Spatz et al., 2023a). They pose a significant threat to seabirds as they prey on their eggs, chicks, and, rarely, on adults, and their eradication or control has been proven to have a positive impact on seabird populations (Howald et al., 2007; Jones et al., 2008). Due to their small size, agility, and ability to hide, rats can be difficult to detect when introduced to new sites. Their rapid reproductive rates and ability to exploit various food sources contribute to their successful establishment and population growth.
Globally, rodents have been eradicated from at least 284 islands, with an ~85% success rate; 387 invasive rodent eradication campaigns were recorded, of which 332 were successful, 35 failed, and 20 were of unknown outcome (Howald et al., 2007). Successful rat eradication/control, mostly through poisoning, has led to an increase in breeding success and breeding pairs, to population growth, the recolonisation of previously abandoned breeding sites and the stabilisation or reduced decline rates of several seabird species. However, not all seabirds, like fulmars, have shown immediate beneficial effects, and some studies have even observed coexistence between rats and seabirds (Jones et al., 2008; Quillfeldt et al., 2008).
Medium-sized mammals, such as cats, mink, and foxes, are more easily detectable but pose greater challenges due to their swimming abilities, long distance movements, and adaptability. Control measures, such as trapping or hunting, are often implemented to manage their populations (Nogales et al., 2004). Medium-sized mammalian predators can prey on seabirds at all stages of their life cycle (Spatz et al., 2023a) and just a small number of individuals can have devastating consequences on seabirds, as they can exterminate entire colonies in a short period.
Cats, in particular, are highly adaptable predators that feed on a variety of animals, even when they have had enough to satiate themselves. They have been associated with significant seabird declines and present additional challenges due to their close association with humans. Of the two studies that examined the effects of cat control, one reported increased survival rates on Black-vented Shearwaters (Keitt and Tershy, 2003), but another did not show a significant beneficial effect on Madeiran Storm Petrels (Ratcliffe et al., 2010). American Mink, being semi-aquatic and a generalist predator, can swim to islets at least 2 km from shore, and even further if linked by an island chain (Ratcliffe et al., 2008). They prey on the eggs and chicks of ground-nesting seabirds. Most of the studies demonstrated beneficial impacts on seabirds following mink control, particularly in terms of breeding success but one study reported no effect, potentially due to the specific focal species, the Great Black-backed Gull (Banks et al., 2008). Foxes, known for their intelligence, can also have devastating effects on seabird colonies, impacting breeding success and adult survival. The only study that focussed on them found that excluding foxes had a positive impact on gulls (Davis et al., 2018).
5.4.5.2 Seabirds’ ecology
The impact of mammal eradication and control efforts is also influenced by the body size and nesting strategy of seabirds. In Table 30, we categorised seabirds based on their nesting ecology into four groups: (1) burrow-nesting birds, including puffins, petrels, and some species of auklets, which excavate burrows in the soil or utilise pre-existing ones for nesting; (2) ground-nesting birds, such as gulls and certain tern species, which lay their eggs on the ground in open areas or rocky outcrops; (3) cliff-nesting birds, including Guillemot, Razorbill, and Kittiwake, which lay their eggs on narrow ledges along steep cliffs or rocky coastal outcrops; and (4) crevice-nesting birds, such as shearwaters, petrels, and Razorbill, which lay their eggs in narrow crevices or cavities commonly found in rocky cliffs or boulder fields (some species use more than one nesting strategy so appear in multiple groups).
These distinct nesting strategies, along with body size, contribute to differing levels of vulnerability to mammalian predators and can affect the success of eradication and control efforts (Lewison et al., 2012). A meta-analysis conducted by Jones et al. (2008) suggested that smaller birds nesting in crevices and burrows, such as storm petrels, are particularly vulnerable to rat predation. Conversely, larger ground-nesting birds, like gulls, exhibit lower vulnerability. Our findings align with these observations.
Most burrow-nesting birds (auks, petrels and shearwaters) were mainly affected by rats. Most of the studies included in this literature demonstrated strong positive effects of eradication efforts on various aspects of their ecology, such as increased breeding success and survival. The only study that did not show a benefit (Lambert et al., 2021), in Manx Shearwaters, was due to the presence of a second rodent predator and not due to the reduced impact of rat removal. Ground-nesting birds, such as gulls, terns, shags, and skuas, also seem to strongly benefit from control and eradication efforts. These species, nesting in open areas, face threats from all invasive mammals, not only from direct predation, but also from trampling and egg displacement when mammals appear at the colony. However, Great Black-backed Gulls were not significantly affected.
Crevice nesters (auks, petrels and shearwaters) were predated by rats, mink and cats. These nests are sheltered and sometimes hard for predators to reach and find, but their eradication still had beneficial effects. Only one record showed no effect, and it was on Madeiran Storm Petrels. Cliff- nesting birds (Kittiwake, auks, Fulmar, and shags) were the least impacted by rat and mink eradication/control efforts. The nests of these species are usually on steep cliffs, inaccessible to predators, so it is not surprising that their eradication did not have a strong effect on them, especially on Fulmar, which have a highly effective defence mechanism of spitting and deterring predators. If a predator managed to infiltrate the colony though, chaos could ensue, leading to the dislodging of eggs and chicks from the cliffs.
Additionally, differences in susceptibility may be attributed to feeding strategies. Burrow-nesting seabirds, like the Atlantic Puffin and petrels, can temporarily leave their young chicks unattended during feeding (even for multiple days in shearwater and petrel species), making them particularly vulnerable to predation at that time. In contrast, ground- and cliff-nesting birds usually always have at least one adult present to protect the eggs and chicks.
5.4.5.2.1 Focal species
Overall, there is evidence that rat eradication in Scotland will likely benefit all the focal seabird species, though to different degrees, whether by increasing their breeding success, pausing, or decelerating decline rates, or by stimulating recolonisations and breeding range expansion. Fox control also seemed to have a strong benefit on gulls, but mink control was beneficial for terns and Razorbill but not for large gulls. Brooke et al. (2018) found faster population growth after eradication efforts among terns and gulls than among other seabird groups, mostly because of their mobility and lack of philopatry to breeding sites.
5.4.5.3 Effects on other species and/or ecosystems
Invasive mammalian predators can have detrimental effects on ecosystems, disrupting the natural balance, impacting native and endemic flora and fauna, and contributing to habitat degradation. Eradicating these predators can allow ecosystems to recover, leading to positive cascading effects on other species and ecological processes (Bried et al., 2009; Jones, 2010). For example, the initially successful eradication of Brown rats from Handa Island not only benefitted seabirds but also resulted in successful breeding for Common Shelduck Tadorna tadorna, Eurasian Oystercatchers Haematopus ostralegus, Common Redshank Tringa totanus and Ringed Plovers Charadrius hiaticula, which was previously not recorded and Pygmy Shrews also appeared to increase on the island (Stoneman and Zonfrillo, 2005). Similarly, mink control efforts in the Finnish Archipelago benefitted seabirds, but also small mammals and amphibians, such as frogs and toads (Banks et al., 2008). While Stanbury et al. (2017) identified 66 species of bird, reptile, amphibian, and mammal that could potentially benefit from invasive mammal eradications across UK islands (and crown dependencies). Jones (2010) showed that soil, plant, and spider-derived nitrogen levels and C:N ratios take mere decades to recover even after centuries of rat invasion. The author also suggests that the recovery of seabird colonies can further speed up the recovery as they provide nutrients that are integral to maintain island biodiversity and ecosystem function.
Nevertheless, eradication/control efforts, if not undertaken in a careful and thoughtful manner, can result in unintended ecological consequences. Many eradication/control methods, such as poison, traps, or guns, can inadvertently harm non-target species, including humans (Appleton et al., 2006; Ratcliffe et al., 2010; Bell et al., 2019). Therefore, it is also important to thoroughly assess the ecological interactions and potential impacts of removing the target mammalian predator, as the absence of such predators may cause changes in prey dynamics and lead to population increases of other predator species, posing a new potential threat to seabirds (Luxmoore et al., 2019; Lambert et al., 2021).
5.4.5.4 External factors that may affect the effectiveness of eradication/control efforts
While eradication and control efforts targeting mammalian predators have shown positive impacts on seabird populations, there are several external factors that can hinder their effectiveness. The effects of climate change, for instance, can lead to habitat loss and a decline in food availability, impeding the recovery of seabird populations (Regehr et al., 2007). For example, in Lewis, terns had low productivity in certain years, despite successful mink control efforts which authors attributed to poor food supply and/or bad weather conditions during that period (Ratcliffe et al., 2006). These results also led authors to believe that in some years, mink predation may not be as harmful, as individuals could be taking eggs and chicks that would otherwise likely have subsequently starved due to poor conditions (Ratcliffe et al., 2006).
The presence of other invasive predator species on the island or region can also undermine eradication efforts. This is of particular relevance to islands in the UK (including Scotland) where the presence of multiple invasive mammalian predators is confirmed or suspected (Stanbury et al., 2017). If only one species is targeted, eliminating that predator can create an ecological vacancy which could allow other invasive species to assume the top predator role, and thus weakening the effect of the eradication (Ballari et al., 2016). This was observed on the Isle of Rum, where localised rat control did not lead to an increase in Manx Shearwater productivity. Authors attributed this to an increase of predation rates by wood mice (Lambert et al., 2021). Furthermore, human activities can disturb nesting behaviours, cause stress, and contribute to declining seabird populations, even in the absence of invasive predators. Therefore, it is crucial to account for these external factors in conjunction with eradication efforts to develop appropriate management efforts for the recovery of seabird populations.
5.4.5.5 Biosecurity measures
The latent risk of mammals reinvading from neighbouring islands or being reintroduced through human activities persists, even after islands have been predator-free for an extended period (Marie et al., 2014). To ensure the success of eradication and control efforts, it is therefore crucial to implement robust biosecurity measures, such as quarantine procedures, monitoring and surveillance systems, and stringent control measures (Holmes et al., 2023). Quarantine and monitoring programmes, such as ‘Biosecurity for Life’, play a crucial role in preventing the introduction (or re-introduction) of species by ensuring restrictions on the movement of people, materials, and animals to and from islands. By closely monitoring these activities, potential threats can be identified and intercepted before they have the chance to gain a foothold in the island ecosystem. These measures are essential for preventing the reintroduction of mammalian predators and the introduction of new invasive species, particularly on islands (Appleton et al., 2006; Banks et al., 2008; Ratcliffe et al., 2008; Phillips, 2010; Marie et al., 2014; Bell et al., 2019; Luxmoore et al., 2019). Neglecting biosecurity efforts can reverse the outcomes of eradication efforts, particularly considering that the positive effects resulting from them may take many years to become apparent. Additionally, measures such as post-eradication habitat restoration programmes and the use of acoustic-playback and decoys can help provide suitable habitat and help attract individuals to further accelerate recolonisations and distribution expansions (Holmes et al., 2023; Spatz et al., 2023a).
5.4.5.6 Social aspects
The eradication and control of mammalian predators, along with the implementation of biosecurity measures, is as dependent on social factors as it is on scientific knowledge and experience (Martin, 2018). As these management strategies become more widely employed, it also comes under increasing public scrutiny (Martin, 2018). It is important to recognise that predator eradication/control efforts, which involve actions such as poisoning, trapping, and shooting animals, can evoke strong emotions and varied opinions within local communities, conservation organisations, and the general public (García-Llorente et al., 2008). This is especially true when working with animals that humans have a greater emotional attachment to, like cats. They are popular as pets and eradication campaigns may face opposition from inhabitants wishing to keep and import domestic animals during and after eradication and because of concerns for their wellbeing (Nogales et al., 2004; Ratcliffe et al., 2010). A feral cat eradication programme, for example, accidentally killed a large proportion of the domestic cat population (Ratcliffe et al., 2010). Failing to account for these opinions and concerns can jeopardise the success of the entire eradication effort, particularly on large, inhabited islands close to the mainland.
Early, inclusive, public consultation and engagement, and transparent communication are therefore essential for building trust and fostering understanding about the need and benefits of such actions (García-Llorente et al., 2008; Crowley et al., 2017). Involving all participants in the decision-making processes and addressing their concerns can help ensure social acceptance and support for this conservation action, both during the eradication process and for the crucial implementation of biosecurity measures (Nogales et al., 2004; Pearson et al., 2019; Holmes et al., 2023).
5.4.5.7 Knowledge gaps and future directions
While the impacts from invasive species are clear, data on invasive mammal presence on islands is still lacking worldwide, potentially underestimating the global threat of invasive species and preventing the identification of areas in need of invasive species management (Spatz et al., 2023a). Even in the UK, where there is generally good monitoring information for certain sites and mammals, there is suspected rather than confirmed presence of predator species (Stanbury et al., 2017). Predator control and prevention in mainland seabird colonies pose unique challenges due to their accessibility to the public. The presence of human activity and coexisting wildlife increases the vulnerability of these colonies to the introduction and spread of invasive species, which further complicates the eradication process. Approaches are less well developed and studied for the control (or ideally eradication) of invasive mammalian predators on mainland or large inhabited islands. For these sites controlling predators is often significantly more challenging than for smaller offshore islands. For the ten UK and Crown Protectorate islands identified as having the greatest potential conservation benefit (this assessment not exclusively for seabirds) from eradication programmes by Stanbury et al. (2017), eradication was only considered realistic for one of these islands as the other nine islands were relatively large with significant human populations. There is, therefore, a need for exploration of control strategies and comprehensive research to enhance our knowledge and inform the development of appropriate management approaches for mainland colonies and for larger inhabited offshore islands.
Achieving successful eradication efforts requires a solid foundation of baseline knowledge, not only about the ecology of seabirds and predators but also about the entire ecosystem. This includes understanding the interactions among multiple co-existing predators and how they regulate each other (Ballari et al., 2016), data which is usually lacking. Therefore, investigating post-eradication ecosystem changes within a whole ecosystem context is vital. It is also important to recognise that the impacts of eradication efforts take time to become visible, particularly when assessing their effects on long-lived species like seabirds. A study on Manx Shearwaters, for example, predicted that during the first six years after eradication the population was going to continue to decline, but after 6–12 years there was going to be a convex growth, and after that an exponential growth, until density dependent limitations cause growth rates to slow (Appleton et al., 2006). Therefore, the success of the eradication effort should be studied over an ecologically appropriate time frame. Given this extended time frame, efforts to accelerate the recovery of seabird populations, like promoting native vegetation or actively restoring seabird populations through activities like translocating chicks or using sounds and decoys to attract prospecting adults should be considered (Benkwitt et al., 2021; Spatz et al., 2023a). Overall, it is crucial to allocate resources and effort towards preventive measures, including the development of monitoring techniques and analytical tools, to predict potential invasion hotspots and prioritise the early detection of invasive species before they pose a significant threat to seabirds and whole ecosystems (Spatz et al., 2023a).
5.4.6 Conclusion
In conclusion, our review provides compelling evidence supporting the effectiveness of mammalian predator eradication as a valuable conservation tool for the protection and enhancement of seabird colonies, particularly for island colonies. The management strategies employed varied depending on the targeted predator, with rats generally being eradicated and larger mammals requiring more focussed control measures. However, the success of these efforts is influenced by multiple factors, including the specific predator, the nesting strategy and body size of seabird species, and various external factors (e.g. distance to mainland and other islands, climate changes, presence of other predator species, terrain, and degree of human presence).
Rats have shown high success rates in eradication campaigns, whereas other medium-sized mammals require more focussed and controlled management. Rats have higher eradication success rates, and mink good control success. Feral cats present unique challenges due to difficulty in gaining community support for lethal control methods. Foxes, on the other hand, exhibit learning behaviours, emphasising the need for adaptable and dynamic efforts for controlling the species. Therefore, eradication and control efforts require adaptive management to accommodate the ecology of the mammalian predators. Most seabird species are expected to show some benefit from mammalian eradication and control efforts, but the degree of the effect will depend on body size and nesting ecology; small burrow and nesting birds are expected to benefit more than large gulls and cliff-nesting birds. Ultimately, rodents mainly affect reproduction (through reduced nest site availability and/or quality or decreased productivity), so their impact tends to be relatively low compared to that of larger invasive species which reduce adult survival (Lewison et al., 2012).
It is crucial to recognise that eradication and control programmes are long-term initiatives, requiring continuous monitoring, preventive measures, and sustained resource investment (Holmes et al., 2023). The timeline for observing positive effects can vary significantly, ranging from short-term to long-term outcomes that may span months or even decades. Additionally, logistical challenges and costs are associated with eradicating mammalian species, particularly in mainland sites and larger islands with higher human population density, making control measures a more feasible management option than eradication. It is essential to consider all predators present on an island during eradication efforts to prevent the rise of alternative predator populations. The success of eradication efforts hinges on the inclusion of local communities throughout the entire process, from the planning stages to the implementation of biosecurity measures to secure the conservation gains and their long-term effectiveness and sustainability. Ultimately, successful eradication or control of invasive species should be viewed as a long-term commitment that requires unwavering vigilance and collaboration among conservation organisations, researchers, and local communities and must sometimes be performed in combination with other conservation actions.
5.5 Ecological feasibility: Avian predator management
5.5.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, the management of avian predators has a low to medium effect on at least one demographic parameter of seabirds, but this strongly depends on the species involved and management type (e.g. diversionary feeding, removal, deterrence, and habitat modification). Considering this conservation action as strategic compensation becomes challenging due to its highly site- and species-specific nature and the overall lack of evidence on their short- and long-term effectiveness. Further complications arise when both the target seabird and predator species are of conservation concern.
5.5.2 Introduction and background
Predation by avian predators, in addition to mammalian predators (as discussed in Ecological feasibility: Mammalian predator eradication and/or management), can exert substantial impacts on some seabird species, particularly those that are ground-nesting and colonial (Roos et al., 2018). Avian predators such as raptors, corvids, and large seabirds like gulls and skuas have been demonstrated to cause detrimental effects on seabird colonies (Parrish and Paine, 1996; Donehower et al., 2007; Hipfner et al., 2012; Smart and Amar, 2018; Anker‐Nilssen et al., 2023; Langlois Lopez et al., 2023; Pollet et al., 2023). These predators can affect seabird populations directly, by targeting eggs and chicks, or indirectly, through disturbance or kleptoparasitism, when an individual steals food or prey from another individual (Finney et al., 2001; Sanz-Aguilar et al., 2009; Perkins et al., 2018). Such disturbances can induce stress in seabirds, leading to behavioural changes that include the reduction of foraging time due to heightened vigilance, an increase in foraging efforts when food is stolen before reaching the chicks, or changes in nesting behaviour as defensive responses. These behavioural changes and predation can consequently affect energetic reserves, breeding success, overall colony structure and distribution, and, ultimately, survival (Gilchrist, 1999; Votier et al., 2004; Oro and Martínez-Abraín, 2007; Perkins et al., 2018; Wilson et al., 2020; Anker‐Nilssen et al., 2023).
Within the UK, numerous avian predators have been observed preying on seabirds. For example, large gulls have been recorded to prey on Roseate Terns in Coquet Island (Alfarwi, 2021), on Common Terns and Black-headed Gulls in Kent (Akers and Allcorn, 2006), and on Arctic Terns in the Farne Islands (Boothby et al., 2019), and are kleptoparasites of Puffins on the Isle of May (Finney et al., 2001; Langlois Lopez et al., 2023). In Great Yarmouth, Kestrels prey on Little Tern chicks (Smart and Amar, 2018) and in Shetland, Great Skuas prey on Kittiwake (Votier et al., 2008). Results from the Breeding Bird Survey (BBS) suggest that populations of half of the common and widespread avian predator species increased significantly between 1995 and 2015 (Roos et al., 2018). While these findings encompass avian predators of all bird species and are not specifically limited to seabirds, the increasing abundance of these predators undoubtedly raises the potential likelihood of intensified predation rates at seabird colonies, and so the potential benefit from predator management.
Understanding the dynamics of prey-predator interactions is particularly essential for developing effective management and conservation strategies, especially when both prey and predator are of conservation concern. There have been many exploratory management actions to reduce avian predation rates in seabird and waterbird colonies, ranging from supplementary and diversionary feeding (Martínez-Abraín and Oro, 2013; Smart and Amar, 2018; Laidlaw et al., 2021) and targeted and untargeted predator nest and/or individual removal (Finney et al., 2001; Akers and Allcorn, 2006; Oro and Martínez-Abraín, 2007; Sanz-Aguilar et al., 2009; Lavers et al., 2010; Paracuellos and Nevado, 2010), to deterrence of avian predators using bioacoustics or physical objects (Boothby et al., 2019; Alfarwi, 2021; Laidlaw et al., 2021), amongst others. The degree of effectiveness of each management action, however, can vary across species, location, and predator-prey interactions. Therefore, targeted management actions tailored to specific contexts are needed and should be carefully designed and implemented considering the ecological dynamics and potential unintended consequences to both prey and predator species.
The aim of this literature review is to identify the main avian predators in Scotland and gather all available evidence of the effect of management actions on seabird species, with a focus on our focal species (i.e. those species most likely to be affected by offshore windfarms). By examining the effectiveness of avian predator management, conservation efforts can be better targeted and prioritised to find the most cost-effective solutions, which may have potential as strategic compensatory measures for the impacts of offshore wind farms on seabirds. Additionally, this review aims to identify which seabird species would benefit most from different avian predator management actions.
5.5.3 Methods
The conservation action, the management of avian predators, involves a set of different management actions and was, therefore, associated with three research questions. The first research question, ‘What is the potential for seabirds to experience increased productivity or survival through avian predator management?’, investigates the impact of avian predator management actions on seabird populations. The second question, ‘Which management action is more effective?’ aims to identify the management action that has the strongest beneficial effect on seabird populations. The third question, ‘For which avian predator is there the most potential for effective management?’, aims to identify the avian species with higher probabilities of management success.
We focussed our search on three management actions deemed relevant for the UK: (1) diversionary feeding, (2) deterrence of avian predators with bioacoustics or physical objects, and (3) targeted nest and/or individual removal or translocation.
A detailed description of the research question, keywords, search strings, and study selection (Figure 19) is provided in section 6 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
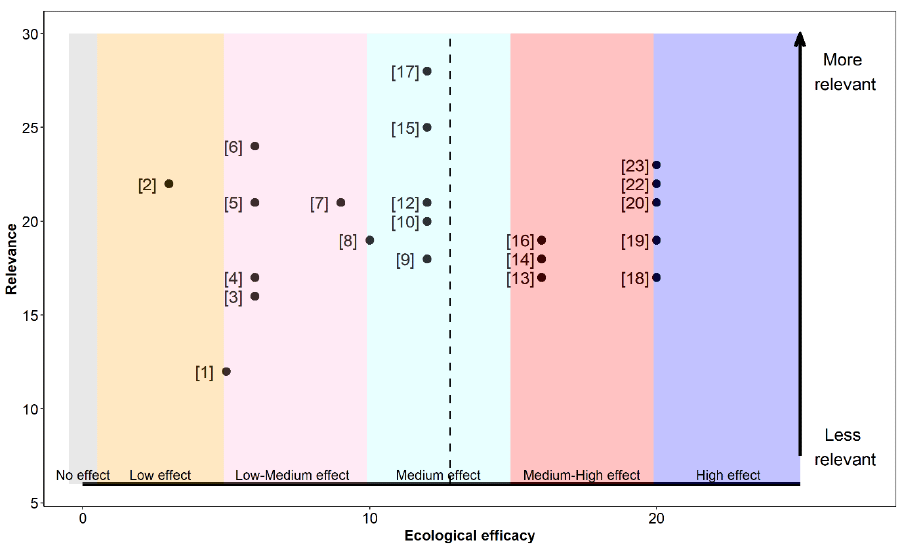
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 11 references were included for the final review.
5.5.4 Results
5.5.4.1 General observations
We identified 11 references that met our criteria of measuring the effect of the three identified avian predator management actions on seabird populations, all published in English. The literature review yielded publications ranging from 1980 to 2021 (Figure 20A). The median publication date was 2001, which also coincided with the only year where more than one study was published. Apart from one short communication, all records were full research articles (Figure 20B). Due to the nature of this conservation action, all studies assessed the impact of at least one avian predator management action. Most studies conducted field experiments to directly test the degree of impact of the management either on seabird or waterbird colonies. Of the assessed studies, 91% focussed on the impact of an avian predator management action on productivity, while 46% studied predation rates, 18% studied survival, 18% studied breeding pairs, and 27% focussed on other parameters such as population size, flush rates, and area occupied for breeding (Figure 20C).
Studies researched the effect of several avian predators on diverse seabird and waterbird species groups (Figure 20D). Out of the studied avian predators, gulls (i.e. Herring Gull, Lesser Black-backed Gull, Great Black-backed Gull, Yellow-legged Gull, Common Gull, and Ring-billed Gull) had the highest number of studies focussed on their management, with eight studies. The second most commonly studied avian predator were raptors (i.e. Kestrel Falco tinnunculus, Red Kite Milvus milvus, and Bald Eagle Haliaeetus leucocephalus), with three studies. One record focussed on Carrion Crows Corvus coronea. Studies focussed on several seabird (or waterbird/wader) species groups (listed from most studied to least): terns (4 studies; i.e. Little Tern Sternula albifrons, Common Tern Sterna hirundo, and Arctic Tern Sterna paradisaea), waders (3 studies; Northern Lapwing Vanellus vanellus, Oystercatcher Haematopus ostralegus, Golden Plover Pluvialis apricaria, Curlew Numenius arquata, Redshank Tringa totanus), auks (2 studies; Atlantic Puffin and Guillemot), and a study on European Storm Petrels Hydrobates pelagicus and one on the Audouin's Gulls Ichthyaetus audouinii. Note that some references studied multiple predators and seabird/waterbird species.
Studies used different methods to manage avian predators, the majority in a non-lethal manner (Figure 20E). Seven studies undertook a removal management action. This type of management included targeted and non-targeted culling of adults, either by shooting, poisoning, or trapping (lethal), and the removal and destruction of nests and eggs to prevent predators from breeding in the area (non-lethal) (Figure 20E). Two studies measured the impact of diversionary feeding of raptors (Kestrel and Red Kite) on Little Terns and Northern Lapwing, respectively. Here they used platforms and/or ground feeding stations to provide 100% of the daily food requirement of the predators for a limited period, mostly during the predator’s chick-rearing period. Two studies undertook habitat modifications, either by providing an artificial habitat that provided protection or by controlling vegetation growth. One study undertook experiments on the effect of placing canes around nests on predation rates. Most studies (64%) were conducted on islands, while the remaining 36% were undertaken on the mainland (Figure 20F). Data was collected from multiple sites across the globe, including the UK, Spain, Canada, and the USA (Figure 20G). Within the UK, six studies were undertaken, three on the mainland and three on islands (Figure 20H). These projects included various sites on the east Coast, such as (from north to south): Kerloch, the Isle of May, Farne Islands, Great Yarmouth, and on the RSPB nature reserve at Otmoor (Figure 20H). Note that locations are grouped per UK-region (northeast Scotland, southeast Scotland, northeast England, and east England), or at a country-level and do not necessarily reflect the exact coordinates of the study sites. Of these, three studies focussed on removal experiments, two on diversionary feeding and one on predator deterrence using canes.
The most frequent data collection methods were observational field data, nest monitoring, pellet collection, chick measurements, and surveys. Most of the authors contributed to one reference.
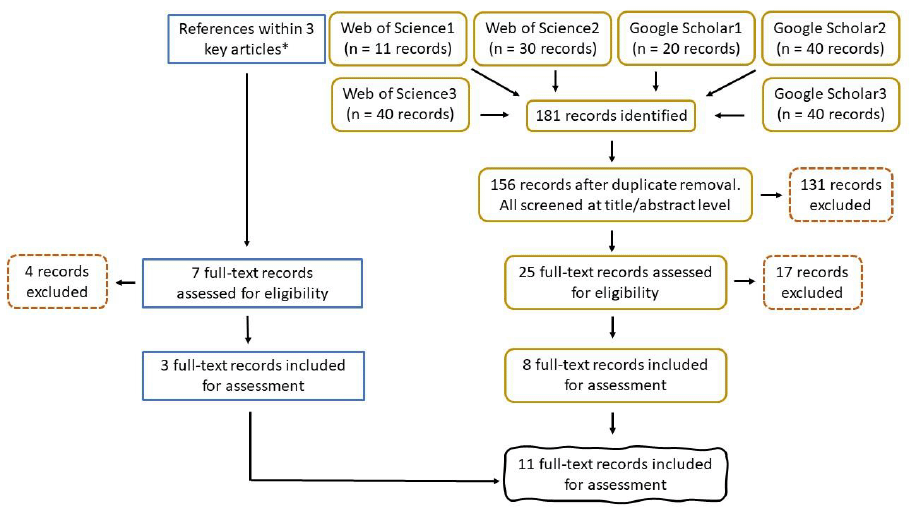
Table 31. Relevance and ecological efficacy scores, as well as the management intervention and its effect on seabirds or waterbirds for each assessed study of the avian predator management review. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of avian predator management, or those with low statistical inference; higher scores reflect studies that found a strong beneficial effect of avian predator management with high statistical inference). REM= Removal, DET = Deterrence, DF = Diversionary feeding, HM = Habitat Modification, SI = Statistical inference, and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
5.5.4.2 Evidence scoring
The relevance, as well as the ecological efficacy scores for all 11 references are shown in Table 31 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Overall, references were of medium-high relevancy for our assessment, with six studies surpassing 20 points. Scores ranged between 15 and 24 out of a maximum possible score of 30, with a median and mean score of 21. For this reason, studies varied in their associated weight, ranging from 0.067 to 0.107 (Table 31). Out of the assessed studies, all but two suggest that managing avian predators has a positive effect on at least one demographic parameter of seabirds or waterbirds.
The ecological efficacy, i.e. the likelihood that the management of avian predators is beneficial on at least one seabird (or waterbird) demographic parameter, varied across studies. Scores ranged between 0 (the management of avian predators does not have an effect on bird colonies) and 25 (the management of avian predators has a strong effect on bird colonies) out of a maximum possible score of 25. The median score was 4, indicating that the effect varied greatly according to each study (Figure 21).
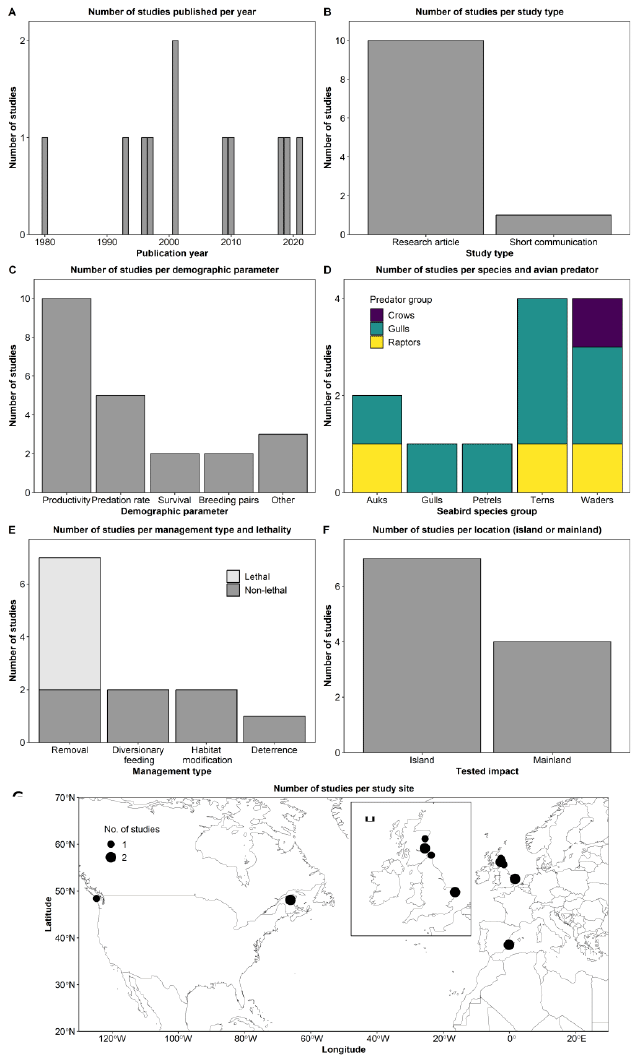
The sum of all final weighted scores is 8 (i.e. the mean of ecological efficacy weighted by relevance), indicating that overall, the evidence suggests that avian predator management will benefit seabirds to a low-medium degree (Figure 21), but this strongly depends on the management type.
5.5.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this conservation action was 8 out of the possible 15 (Table 32), suggesting that we have a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of the management of avian predators.
Table 32. Assessment of the confidence in the evidence for management of avian predators.
This table can be viewed in Excel format.
5.5.4.4 Main findings: the effect of avian predator management
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, the management of avian predators has a low-medium effect on at least one demographic parameter of seabirds, but this strongly depends on the management type.
The key findings from the most relevant studies are summarised in Table 33. Overall, the effect of avian predator management had, to some degree, a positive effect on at least one demographic parameter of colonial birds, including seabird and waterbirds. The degree of this effect, however, varied strongly on the management type (Table 34).
Two studies researched the impact of diversionary feeding (DF) of raptors on the productivity of two ground nesting species, the Little Tern (Smart and Amar, 2018) and Northern Lapwing (Mason et al., 2021) in eastern England. Both studies compared predation rates and productivity during DF and non-DF years. Smart and Amar (2018) found that Kestrel predation rates were 47% lower (or 88% during additional intensive monitoring) and productivity of Little Terns doubled during DF years, from a mean of 0.42 fledged chicks/pair in non-DF years to 0.88 in DF years, though the difference was not statistically significant. Mason et al. (2021) found extremely similar results but when studying diversionary feeding of Red Kites. Their results also show lower predation rates during DF years and a mean increase in lapwing productivity, from a mean of 0.47 fledged chicks/pair in non-DF years to 1.02 in DF years.
Table 33. Summary findings of the five most relevant studies on the effect of avian predator management on bird colonies, in order of decreasing relevance.
This table can be viewed in Excel format.
Table 34. Key findings of the studies for each seabird or waterbird species within the avian predator management review. Focal species are highlighted in italics. The effect is colour-coded as follows: red or ‘=’ = no effect, white or ‘+’ = small effect but not enough to reflect a population increase, and blue or ‘++’ = effect that led to a population increase (e.g. measured increase in productivity or population growth).
This table can be viewed in Excel format.
Removal techniques and their effect on seabirds and waterbirds varied strongly among the seven studies that tested this type of management. Techniques for gull control ranged from targeted and long-term culling to nest and/or first clutch removal, as well as egg pricking (piercing a small hole in the shell to prevent hatching). One study undertook this jointly on both gulls and crows (Parr, 1993). Targeted culling of gulls aims to remove specific individual gulls that are thought to contribute the highest proportion of predation of other species. Both studies that undertook this found significant increases in survival and/or breeding success of seabirds (Guillemette and Brousseau, 2001; Sanz-Aguilar et al., 2009). Sanz-Aguilar et al. (2009) observed that the removal of six specialised Yellow-Legged Gulls and ten additional individuals nesting in the proximity of a nesting site (a cave) used by European Storm Petrels led to a mean increase in adult survival probability from 0.75 to 0.89 (95%CI = 0.82–0.94) and to an increase in breeding success from 0.50 to 0.66. The specialist gulls were mainly territorial males and represented a small proportion of the population (ca. 1%). Guillemette and Brousseau (2001) similarly found that removing four male Herring Gulls and one Great Black-backed Gull decreased predation rates on Common Terns and increased their fledging success from 0 to 16% of chicks that hatched surviving to fledging. One individual gull accounted for 85% of all successful predation attempts made during the baseline period.
Non-targeted culling programmes seem to have a mixed effect on colonial birds. In Spain, a Yellow-legged Gull culling programme significantly increased the number of breeding pairs, area occupied and breeding success of the Audouin’s Gull population (Paracuellos and Nevado, 2010). On the Isle of May, however, gull culling increased the number of breeding pairs of Oystercatchers but did not significantly improve their breeding success (Harris and Wanless, 1997b). In Kerloch moor, the removal of both Carrion Crows and Common Gulls did not have an effect on waders (Parr, 1993). Although the numbers of crows and gulls were significantly reduced, no young Golden Plover hatched, and breeding numbers continued to decline. Hatching success of Oystercatchers also did not increase, and although Lapwing, Curlew, and Redshank had higher hatching success, their breeding numbers did not increase. Authors believe that these results can be explained by the rising nest predation from foxes, which may have masked any benefits from the removal of the other predators.
Removal does not only include targeting adults but includes removing nests and/or egg clutches and preventing individuals to breed at sites close to other species. On the Isle of May, Finney et al. (2001) compared kleptoparasitism rates (when an individual ‘steals’ food from another) and the reproductive performance of Puffin. Specifically, the compared these variables among Puffins breeding in gull-occupied habitats with those nesting in areas where gull breeding was delayed through the removal of the first clutch. Overall, kleptoparasitism was lower within gull-free plots. However, the breeding performance (measured with breeding success, hatching date, peak weight, fledging period, fledging weight) was the same in all treatments. In addition to habitat modification, Morris et al. (1980) showed that the removal of Yellow-legged Gulls nests increased the breeding success of Common Terns.
Deterrence using a physical object was only measured in one study in the Farne Islands. Boothby et al. (2019) tested a new technique, using bamboo canes of 1.5 m in length at an approximate angle of 70° to the ground, to deter gulls from Arctic Tern nests. They measured predation rates and predation success using three treatments which included a control site, a site with a low density of canes, and a site with a high-density of canes. Authors found that gulls had lower predation rates (up to half) in caned sites compared to the control sites but that if an attempt took place, then canes did not affect whether it would be successful.
Two studies explored the effect of habitat modification on seabirds. The first, explored the effect of creating temporary silk-enhanced artificial trees (‘silk forests’) to protect Guillemots from Bald Eagles in the USA (Parrish and Paine, 1996). This predator represents more of a perceived rather than an actual threat, as it induces the desertion of Guillemots' nests on the cliffs, without necessarily consuming the eggs/chicks. However, during this period crows and gulls can predate the unprotected nests. In this experiment, authors showed that Guillemots colonised the artificial habitat and even showed twice the density of eggs and had lower flush rates during eagle attacks compared to those from the control sites. However, productivity did not increase as no chicks hatched on any site. Authors believe that the artificial habitat can increase productivity, but at a larger scale. The second study combined gull-removal techniques and the control of vegetation growth to enhance Common Tern productivity in Canada (Morris et al., 1980). They showed that Common Terns had higher breeding success rates, both of eggs hatched/eggs laid and chicks fledged/eggs hatched, during the year with management actions than during the control year, but only when comparing early nesters (the managed year had a longer breeding season so many pairs relayed. Overall, late nesters had lower success than early breeders).
5.5.4.5 Biases or Conflict of interest
The authors did not disclose any biases or conflicts of interest, and our investigation did not reveal any such concerns.
5.5.5 Discussion
Evidence suggests that, to some degree, the management of avian predators has a beneficial effect on at least one demographic parameter of seabird populations during the breeding season, albeit to varying degrees. The effectiveness of such management actions is influenced by the avian predator involved, the nesting ecology of the target seabird species, and most importantly, the type of management which, in turn, was contingent upon factors such as the specific predator species, prey species, and geographical location. The management types explored during this literature review encompassed temporary diversionary feeding of raptors, the use of gull-deterrent objects, the removal of gulls and their nests and habitat modifications, all of which are discussed in detail below.
5.5.5.1 Diversionary feeding
Diversionary feeding (DF) is a non-lethal, temporary method aimed at diverting the activity or behaviour of predator species away from actions that negatively impact a particular target population (Kubasiewicz et al., 2016). Distinct from supplementary feeding, which aims to improve the population viability or density of a particular species or population, DF does not seek to increase the density of the population (Kubasiewicz et al., 2016). Studies by Smart and Amar (2018) and Mason et al. (2021) demonstrated the efficacy of DF of raptors in reducing predation rates and doubling the productivity of Little Terns and Northern Lapwings, respectively. During DF years, Kestrels preferred to hunt wild mammals over Little Terns, suggesting that terns may be considered a riskier prey type and are only targeted when they are abundant or when mammal prey is scarce (Smart and Amar, 2018). These findings suggest that DF of certain raptors, such as Kestrels and Red Kites, can be a highly effective predation management tool that enhances productivity of ground-nesting colonial species that could consequently lead to population growths. During this review, no evidence was found regarding its applicability and efficacy on predator gulls and/or its potential for protecting non-ground nesting species (e.g. crevice and cliff nesters, such as auks).
DF proves particularly useful when both prey and predator species are of conservation concern, as it reduces the motivation for a predator to hunt natural prey without compromising the needs of either species (Smart and Amar, 2018). However, it can also cause unintended consequences (Martínez-Abraín and Oro, 2013). DF has the potential to attract or recruit new individuals, increasing predator abundance and density and thereby increasing predation rates (Cortés-Avizanda et al., 2016; Mason et al., 2021). It can also create dependence on human-provided food that may not be sustained once this food source is removed or depleted (Kubasiewicz et al., 2016), and, if it attracts a high density of individuals, it can increase the risk of disease transmission. Precautionary measures can, however, be taken to mitigate these risks. Mason et al. (2021), for example, propose implementing DF for a short period, beginning after most raptor pairs have chosen their nest sites and invested energy in egg laying and incubation and in a site close to where the highest prey chick predation occurs. Another solution could be to use DF during periods with peak predation rates (e.g. mid-breeding season). This timing is likely to reduce predation in the short-term. To reduce costs and prevent predators from becoming habituated to DF, authors also suggest implementing DF in alternating years.
When designing a successful and appropriate DF activity, Smart and Amar (2018) suggest considering several factors: the choice of food, feeding location, timing and method; determining the appropriate food quantity to provide; minimising food intake by non-target species; understanding the specific needs and preferences of the target species; maintaining consistency in the timing and frequency of the feeding; and aiming to provide 100% of their dietary requirements. However, it is important to note that many more studies are needed in order to properly understand the effectiveness of this management as a long-term conservation tool on several species (Kubasiewicz et al., 2016).
5.5.5.2 Removal
The predominant management action identified in this literature review involves the targeted or non-targeted removal of predators’ nests, eggs, chicks and/or breeding individuals. The focus of such actions has primarily been directed towards large gulls, likely due to their population increase and distribution expansion resulting from fishery discards and landfill sites. Numerous long-term culling programmes have been implemented worldwide, aiming to reduce predator presence in specific sites. These programmes are generally conducted at the local population level and consist of the systematic lethal removal of large numbers of eggs, chicks, or breeding adults. This action assumes that all individuals contribute equally to, or are equally responsible for predation attempts, and targeting them accordingly.
Nonetheless, evidence suggests that the long-term success of gull control programmes is relatively low, as there is no guarantee of an increase in population size or breeding success, even when substantial efforts are undertaken, particularly for waders (Parr, 1993; Harris and Wanless, 1997b; Oro and Martínez-Abraín, 2007). The review identified only one successful event where non-targeted culling of gulls improved the breeding success of a smaller, threated seabird species (Paracuellos and Nevado, 2010). This occurred at a small, isolated colony, which authors believe was the reason of the success. However, Paracuellos and Nevado (2010) suggest that this action should not be seen as a viable long-term solution. Instead, they advocate for addressing the underlying factors that restrict predator populations as a more effective strategy.
Targeted (or selective) removal, on the other hand, has shown more significant beneficial effects on breeding success and survival in seabirds like terns and petrels (Guillemette and Brousseau, 2001; Sanz-Aguilar et al., 2009; Scopel and Diamond, 2017). This approach not only reduces overall project costs but also has a lesser impact on the predator population. Sanz-Aguilar et al. (2009) observed that specialist gulls with higher predation attempts were mainly territorial males and represented ca. 1% of the populations. These individuals can be individually identified and removed at the beginning of the prey species’ breeding period due to their territorial behaviour, but this does not apply to juveniles or non-breeding individuals (Guillemette and Brousseau, 2001). It is important to note that even in cases where dominant individuals are successfully eliminated, continuous monitoring and culling programs remain necessary, as successive predatory gulls could replace one another when dominant individuals are removed (Guillemette and Brousseau, 2001; Votier et al., 2004).
If an avian predator, regardless of the nesting ecology of its prey, were to be identified and could be successfully removed, then, in theory, this management strategy should be effective. Nest removal and egg pricking could be additional activities that may reduce the predators breeding success (Morris et al., 1980; Smith and Carlile, 1993), though this did not work for Puffins (Finney et al., 2001). These actions however mean that adults remain alive at colonies and may continue preying on individuals.
5.5.5.3 Deterrence
The use of deterrent objects, including physical, acoustic, or chemical means, has been explored as a strategy to deter birds in areas such as landfills, airports, and agricultural sites e.g. (Baxter and Robinson, 2007; Cook et al., 2008; Soldatini et al., 2008; Peterson and Colwell, 2014). However, their application at seabird colonies remains relatively unexplored. A pilot study conducted by Boothby et al. (2019) examined the use of bamboo canes as a physical deterrent at a tern colony and found a reduction in predation attempts but no significant improvement in breeding success. It is important to note that this study had limited sample sizes, and further research with larger sample sizes over multiple breeding seasons is necessary to ascertain the potential effectiveness of such deterrent objects.
The overall aim of deterrent objects is to alter an avian predators’ behaviour and discourage their presence in certain areas. Physical deterrent objects encompass a range of visual stimuli designed to elicit an aversive response in large birds which could include scarecrows, decoys, reflective materials, bamboo canes, or lasers (Peterson and Colwell, 2014; Alfarwi, 2021). Acoustic deterrents utilise various auditory signals, including distress calls, predator vocalisations, or avian alarm sounds, to repel birds (Thieriot et al., 2015). Chemical deterrents involve the use of substances that produce aversive or repellent effects. Understanding which deterrent would work at seabird colonies should be further tested and at this point it is difficult to conclude anything from the lack of evidence found in the literature review. Most likely, however, the degree of effectiveness and implementation must be highly species- and colony- specific. Important to consider that predators could habituate to certain deterrents over time, reducing their overall efficacy.
5.5.5.4 Habitat modification
An additional management approach that had not been initially considered was habitat modification, which emerged as a frequently employed strategy and was sometimes used as a complementary action. The results from habitat modification studies showed mixed effects but were overall beneficial. This approach encompassed diverse methods. One study, for example, used artificial ‘silk forests’, a temporary and reversible new habitat to reduce the threat perception of Common Guillemots by Bald Eagles (Parrish and Paine, 1996). The temporary nature of this method avoids any impact on Eagles and allows the habitat to revert to its original state after its removal, at the end of the breeding season. This type of action can be used as an interim solution while longer-term conservation measures take effect (Parrish and Paine, 1996).
Another technique of habitat modification involves managing vegetation growth in accordance with the nesting preferences of seabirds. Terns, for example, prefer nesting in areas with low vegetation as denser and taller vegetation can reduce visibility and increase the risk of entanglement (Morris et al., 1980). What vegetation or general habitat modifications should be undertaken should strongly depend on the specific preferences of the target species. Providing an appropriate level of vegetation in front or around Puffin burrows, for example, could aid in protecting pufflings and potentially reducing levels of kleptoparasitism from gulls. Modifying habitat of cliff-nesting species may present logistical challenges.
5.5.5.5 Knowledge gaps and future directions
It is evident from this review that evidence is lacking for many management actions, indicating a need for further research. To obtain a more comprehensive understanding of which actions are effective for different seabird species, future studies should focus on filling these knowledge gaps.
Although we included information on the effects of avian predator management on colonial waterbirds, our literature search was focussed on seabirds, and we are confident that much information on this was overlooked. Conducting a review that includes similar colonial species, such as waders, could provide valuable insights into the effectiveness of conservation actions. Results from this review could increase our understanding of the applicability and potential transferability of management strategies to different seabird species.
Furthermore, conducting reviews specifically focussing on the deterrence of target avian predators in non-seabird colony sites such as landfills and airports, could offer insight into the mechanisms that effectively deter these predators. For example, falconry, robots, pyrotechnics, and the playback of distressed calls have shown to successfully deter gulls from certain areas (Baxter and Allan, 2006; Baxter and Robinson, 2007; Cook et al., 2008; Soldatini et al., 2008; Thieriot et al., 2015; Storms et al., 2022). Translocation, the capture, transport and release of individuals from one location to another, could also be further explored as a management option (Ackerman et al., 2014), though this would be species-specific and would require a high cost. Understanding which deterrent methods are successful in repelling focal predator species could help identify potential strategies that could be adapted for use at seabird colonies, without negatively impacting the seabirds themselves.
5.5.6 Conclusion
Top-down effects from avian predators can be significant regulators of seabird populations (Anker‐Nilssen et al., 2023). Overall, our findings indicate that, to some degree, the management of avian predators can have a beneficial effect on seabird populations during the breeding season, albeit to varying degrees. The effectiveness of such management actions is heavily influenced by factors such as the avian predator involved, the nesting ecology of the target seabird species, and most importantly, the management action (e.g. diversionary feeding, removal, deterrence, and habitat modification). Each management action possesses inherent advantages and disadvantages that require careful consideration during the planning and implementation stages and should always be tailored to the specific seabird species and avian predator in question. Considering this conservation action as strategic compensation becomes challenging due to its highly site- and species-specific nature and further complications arise when both the target seabird and predator species are of conservation concern.
Most of these management actions can be undertaken at small scales and during short periods of time, often aligned with the breeding season of the target seabird species, at relatively lower costs compared to other of the conservation actions explored in this report. However, the current lack of evidence on the short- and long-term effectiveness of avian predator management as a conservation tool requires further research (Kubasiewicz et al., 2016).
5.6 Ecological feasibility: Reduction of disturbance (at colony)
5.6.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, management measures of disturbance at and around colonies have a low to medium effect on at least one demographic parameter of seabirds. We suggest that reducing disturbance at colonies could be beneficial for seabirds. However, due to uncertainties regarding population level benefits, further research is needed before being considered for use as strategic compensation.
5.6.2 Introduction and background
Following Coetzee and Chown (2016) and Allbrook and Quinn (2020), human disturbance to wildlife refers to any activity, event, or action conducted by humans that induce physical, behavioural, or physiological changes in individuals, potentially leading to short- or long-term stress or fitness responses. While human disturbance affects most coastal seabird species, its impact is relatively lower compared to other threats like invasive alien species, bycatch, and climate change (Croxall et al., 2012; Dias et al., 2019). Nonetheless, nesting seabirds, particularly those nesting in accessible locations, are highly vulnerable to human disturbance as they are bound to their nests and are exposed to disturbance occurring from water, land, and more recently, air. Factors contributing to disturbance at colonies include tourism, recreation, photography, research, pedestrians, pets, and vehicles on land and at sea. More recently, the increasing use of Unmanned Aerial Vehicles (hereafter UAVs) in ecological research, photography, and recreation (Brisson-Curadeau et al., 2017) has introduced a new aerial disturbance.
There is extensive evidence showing that human disturbance has negative impacts on nesting seabirds. Such disturbance can result in increased nest abandonment, extended foraging time, changes in foraging locations at times abandoning preferred areas, changes in behavioural responses (e.g. resting, vigilance, flushing, agitation), shifts in habitat use, higher energy expenditure, reduced incubation time, increased predation risk, site abandonment, reduced parental investment and, in some cases, direct mortality (Chardine and Mendenhall, 1998; Carney and Sydeman, 1999; Blanc et al., 2006; Price, 2008; Watson et al., 2014; Allbrook and Quinn, 2020). Some consequences are more challenging to detect as they are not visible, as disturbance can cause physiological impacts such increased stress and cardiac rhythm, and reduced immunity (Ellenberg et al., 2006). These effects can subsequently influence body condition, reproductive success, and overall fitness, with potential population-level implications such as changes in population trends or permanent colony abandonment (Carney and Sydeman, 1999; Blanc et al., 2006; Allbrook and Quinn, 2020).
To mitigate the impact of human disturbances on seabirds, a range of management measures have been proposed at seabird colonies (e.g. visitor regulations, use of warning signs, setting speed limits, and regulating the use of UAVs). Understanding the effectiveness of such management measures and their specific impacts on seabird physiology, demography, and behaviour, is a vital step in designing and implementing appropriate management measures to enhance seabird conservation. Understanding why and how birds respond to disturbance may give insights into how conservation managers may minimise the impact of human disturbance at seabird colonies.
Various seabird species exhibit varying degrees of sensitivity to human disturbance, with some species more susceptible than others (Yorio et al., 2001; Blumstein et al., 2005; Price, 2008; Chatwin et al., 2013; Bishop et al., 2022). While some species can adapt and minimise the effects resulting from disturbance, others may remain highly sensitive, making it essential to tailor conservation and management strategies based on the specific needs of each species. The aim of this review is to gather evidence on the effect of management measures to reduce disturbance at seabird colonies, with a particular focus on our focal species and to identify which seabird species would benefit most from different disturbance management measures.
5.6.3 Methods
This conservation action was associated with two research questions. The first research question, ‘What are the potential population level benefits from reducing on-land and coastal disturbance at seabird colonies?’, investigates the impact of disturbance management measures on seabirds when present at the colonies. The second question, ‘What types of disturbance management measures will provide the strongest benefit?’ aims to identify the measure/s that could have the strongest beneficial effect on seabirds. Note that this review focusses solely on mostly recreational disturbance occurring during the breeding season, and management actions will focus on reducing disturbance directly on land, or at sea, but close to shore, enough to disturb birds breeding on land.
A detailed description of the research question, keywords, search strings, and study selection (Figure 22) is provided in section 7 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 10 references were included for the final review.
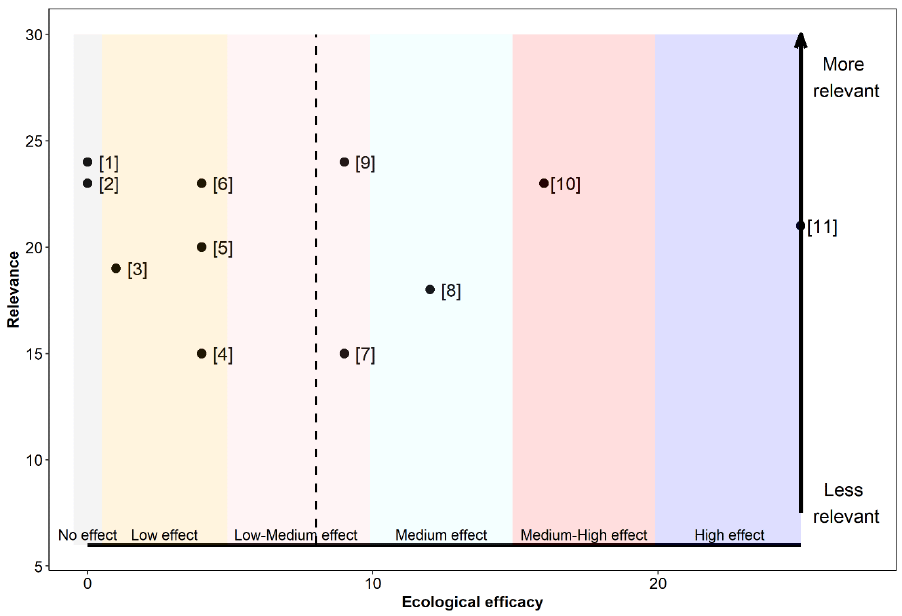
5.6.4 Results
5.6.4.1 General observations
We identified ten references that met our criteria of measuring the effect of disturbance at or close to seabird colonies. Nine of these studies were published in English, while one was published in Slovenian. The literature review yielded publications ranging from 2002 to 2022 (Figure 23A), with a median publication date of 2012. All records were research articles. Studies mostly conducted behavioural observations and experiments to test the degree of impact that land or sea disturbance had on seabirds. Half of the assessed studies focussed on the impact of disturbance on breeding success and the other half on behavioural responses, such as flushing rates or signs of agitation (Figure 23B). It is important to note that while behavioural responses are not demographic parameters per se, they can serve as potential drivers of demographic parameters. However, it is highly challenging to understand how changes in behaviour reflect on changes in population dynamics.
Seven studies conducted field experiments to directly test the degree of impact of certain management measures on seabirds at colonies, while the remaining three tested it indirectly (Figure 23C). Six studies researched visitor management at colonies (i.e. set-back distances, signage, and restriction of number of visitors), one study focussed on the effect of unmanned aerial vehicles (UAVs), and the rest tested measures relating to boat and kayak management close to colonies (i.e. set-back distances at sea and speed limits of vessels; Figure 23D). Two studies were conducted at St Abbs Head, Scotland, while the others were undertaken outside of the UK, in Canada, the USA, Ireland, Portugal, Slovenia, and Mexico (Figure 23E).
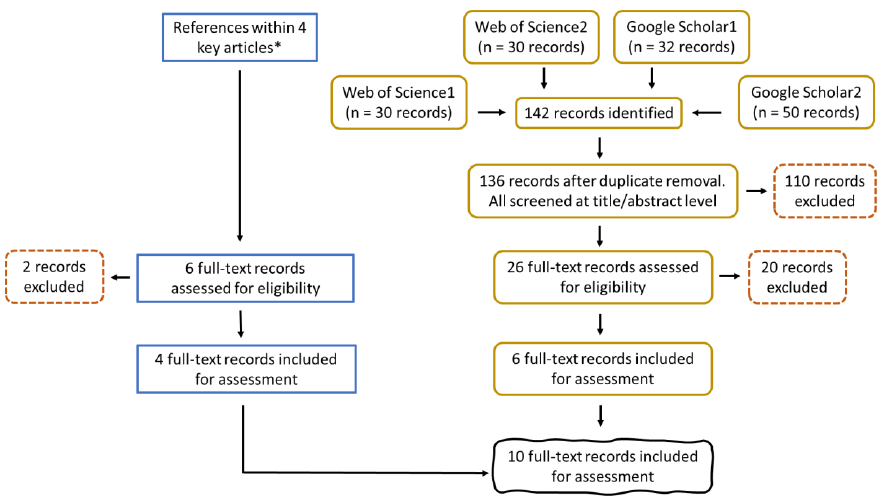
Data was collected from multiple species, mostly on auks (Guillemot, Pigeon Guillemot, Black Guillemot, Cassin’s Auklet), followed by gulls (Kittiwake, Black-headed Gull, Glaucous-winged Gull), terns (e.g. Common Tern, Roseate Tern, Little Tern), cormorants (Brandt’s Cormorant, Double- crested Cormorant, Pelagic Cormorant), and Gannet. Some references researched multiple species.
5.6.4.2 Evidence scoring
The relevance, as well as the ecological efficacy scores for all ten references are shown in Table 35 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Overall, references were of low-medium relevancy for our assessment, with only two studies surpassing 20 points. Scores ranged between 15 and 23 out of a maximum possible score of 30, with a median score of 19.5 and a mean score of 19. Studies varied in their associated weight, ranging from 0.079 to 0.105 (Table 35). Out of the assessed studies, all suggested that measures to reduce disturbance at colonies could, potentially, have a positive effect on reducing unnecessary behavioural changes and on at least one demographic parameter of seabirds. Bishop et al. (2022), however, found that the effect of UAVs was species-dependent and, therefore, disturbance management measures may be more beneficial for some species over others.
Table 35. Relevance and ecological efficacy scores, as well as the type of regulation and whether it applies on land, at sea, or by air, and its effect on seabirds for each assessed study of the disturbance management at and around the colony review. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of the disturbance management measure, or those with low statistical inference; higher scores reflect studies that found a strong beneficial effect of the management measure and high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
The ecological efficacy, i.e. the likelihood that measures to manage disturbance on land, at sea, or airborne (e.g. UAV) around colonies is beneficial on at least one seabird demographic parameter, varied across studies. Overall, ecological efficacy scores were low, ranging from 3 (management measures at colonies had minimal effect on seabirds) to 12 (management measures at colonies had a medium effect on seabirds) out of a maximum possible score of 25. The median score was 3.5 (Figure 24). The lower scores observed can be attributed to the fact that the studies primarily focussed on measuring behavioural aspects rather than directly measuring the effects of management measures on demographic parameters (e.g. it is unclear how and to what degree higher flushing rates affect productivity and/or survival).
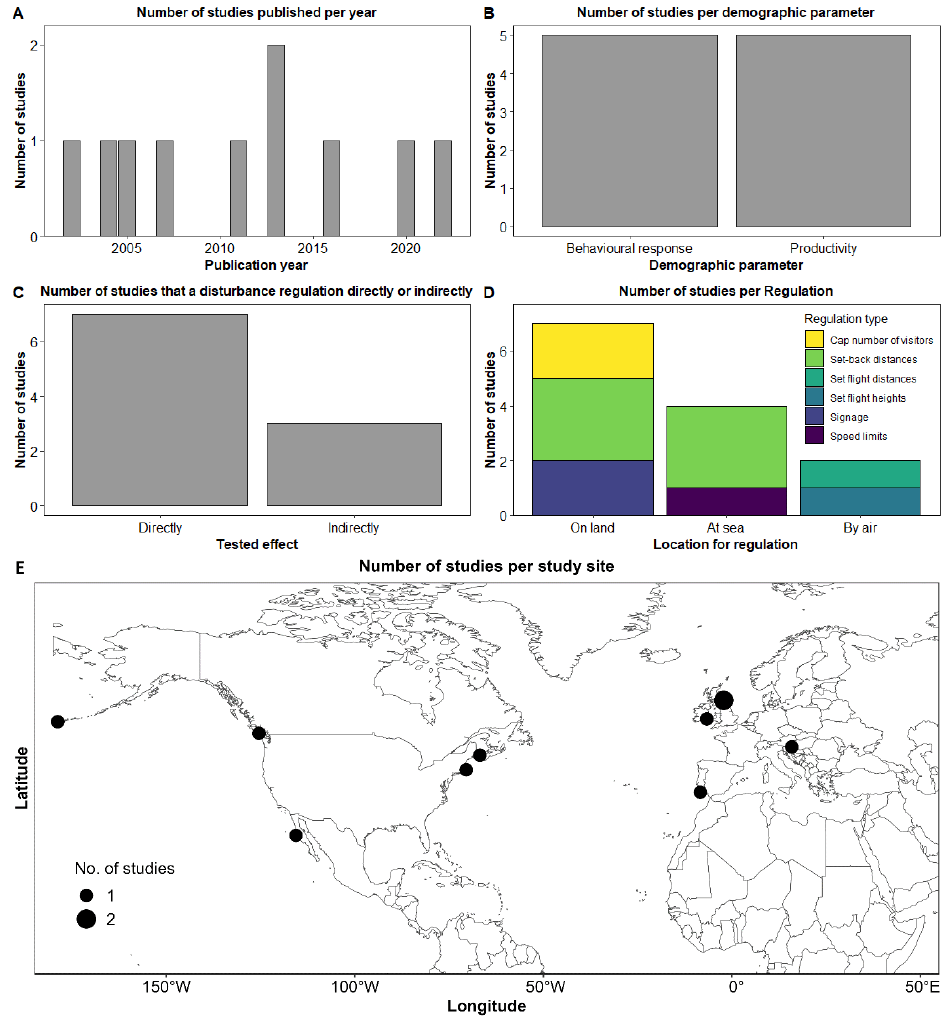
The sum of all final weighted scores is 5.2 (i.e. the mean of ecological efficacy weighted by relevance), indicating that overall, the evidence suggests that management measures at-colony to reduce disturbance will benefit seabirds to a low-medium degree (Figure 24).
5.6.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this measure was 9 out of the possible 15 (Table 36), suggesting that we have a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the ecological efficacy of management measures to reduce disturbance at the colony.
Table 36. Assessment of the confidence in the evidence for management of at-sea disturbance.
This table can be viewed in Excel format.
5.6.4.4 Main findings: the effect of reduction of disturbance at the colony
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, management measures of disturbance at and around colonies has a low-medium effect on at least one demographic parameter of seabirds. This, however, needs to be treated with caution given that half of the studies recorded changes in behaviour responses towards disturbance, making it challenging to accurately quantify their impacts on demographic parameters (e.g. productivity and survival).
The key findings from the most relevant studies are summarised in Table 37. Most studies directly tested the effectiveness of management measures by recording the distance at which seabirds exhibited behavioural distress or flushing, as well as differences in productivity in relation to proximity to pathways. Indirect assessments were conducted by examining human responses (e.g. how close they got to nests) when signs were present, and relating it to differences in breeding success of nests at different distances of pathways.
On land, the management measures that were tested were setting a fixed set-back distance of visitors to nests, managing the number of visitors, and the use of signage to reduce visitor effects on nest disturbance. The most common on-land management measure involved estimating set-back distances and regulating the number of visitors allowed at a site. Most studies compared productivity between highly disturbed plots (i.e. plots closest to paths, which also received more visitors) and less-disturbed plots (i.e. plots farther away from paths and visitors).
Table 37. Summary findings of the five most relevant studies on the effect of management measures to reduce at-colony disturbance on seabirds, in order of decreasing relevance.
This table can be viewed in Excel format.
Overall, nests located closer to paths had lower breeding success (Beale and Monaghan, 2004; Albores-Barajas and Soldatini, 2011; Althouse et al., 2016; Allbrook and Quinn, 2020). In St Abbs Head, Scotland, Kittiwake, in particular, seemed to be more susceptible to human disturbance than Guillemot (Beale and Monaghan, 2004; Beale and Monaghan, 2005). In Mexico, Cassin’s Auklets, a burrowing auk, located within 30 m of paths had significantly lower breeding success (~0.45) compared to those located more than 50 m from paths, despite all having similar hatching rates (Albores-Barajas and Soldatini, 2011). Common Tern and Roseate Tern in Massachusetts, USA, exhibited higher flush rates in response to pedestrians than to researchers. The predicted flush probability increased from 0 to 6% when researchers approached within 50 m. However for pedestrians, the predicted flush probability increased from 6% to ~35% (Althouse et al., 2016). The authors suggest that the speed, angle, and consistency of the approach may explain the differential responses of terns to pedestrians and researchers.
Two studies examined the effectiveness of signs in reducing human disturbance at seabird colonies. In Ireland, Allbrook and Quinn (2020) found that regulatory signage significantly influenced visitor behaviour, leading to a reduction in visitor proximity to Gannet nests when signs were present compared to periods without signs. This decrease in visitor proximity resulted in fewer birds displaced from their nests. Notably, photographers were the only visitor group that did not comply with the signage. Similarly, in Portugal, Medeiros et al. (2007) discovered that the presence of protective measures (warning signs and wardening) was the most important predictor of Little Tern nesting success. Birds were up to 34 times more likely to succeed when these measures were in place. Authors also found that earlier breeders were the most to benefit from the protective measures.
At-sea studies focussed on the behavioural response of seabirds to approaching vessels, primarily motorboats and kayaks. All three studies estimated the distance at which birds flushed in response to vessels. In Vancouver Island, roosting seabirds were generally more sensitive to boat disturbances than nesting birds (Chatwin et al., 2013). When birds were nesting and vessels approached at 30 m, Glaucous-winged Gulls showed a lower probability of agitation (8.3%), followed by Pigeon Guillemot (11.2%), Pelagic Cormorant (15.5%) and Double-crested Cormorant (21.6%). No agitation was recorded for any species at 70 m. Authors recommend setting a single set-back distance, which, at this site, is 50 m. Similarly, the flushing probability of Black Guillemot in Canada, decreased as boat approach distance increased, with the greatest flushing probability associated with fast boats and closer approach distances (Ronconi and Clair, 2002). In Slovenia, Black-headed Gulls flushed when motorboats approached within 67 m, returning after 11 minutes, and Common Tern flushed when the boats were at 73 m, returning within 5 minutes (Vogrin, 2013).
Only one study tested the impact of drones or UAVs at seabird colonies. Bishop et al. (2022) tested the effect of two models of UAVs flown at different speeds and distances on the behaviour of Guillemot, Kittiwake and Cormorants during the incubation and chick-rearing period in Alaska. Neither Cormorants nor Guillemot exhibited signs of flushing or changes in individual behaviour during UAV flights. In contrast, Kittiwake did flush, and there was individual variation in their activity budgets, although most of these birds were not incubating or brooding. At both the colony and individual level, there was a slightly greater behavioural response to smaller UAVs and closer approaches.
5.6.4.5 Biases or Conflict of interests
The authors did not disclose any biases or conflicts of interest, and our investigation did not reveal any such concerns.
5.6.5 Discussion
Our review highlighted that human disturbance in the vicinity of seabird breeding colonies, both nearshore and on land, can originate from various activities. These disturbances can have a wide range of impacts, ranging from minor to severe. Overall, we concluded that reducing disturbance could have a positive effect on seabird populations, however, the evidence available limits our confidence in this assessment with most studies measuring behavioural responses (e.g. avoiding flushing) rather than measures of population level impact (e.g. breeding success).
The level and types of disturbance for a given seabird colony will be site-specific and will depend on the range of activities present. Disturbance can originate from: land-based activities (e.g. humans and pets); sea-based activities (e.g. recreational watercraft); and aerial disturbances (e.g. UAVs such as drones). As such, managing disturbance will largely require site-specific measures, though there can be benefit in these being underpinned by national level regulations and guidance.
Regarding disturbance originating on land, largely from visitors at seabird colonies, the main actions identified include set-back distances, controlling the number of visitors, and signage. Signage alone has been shown to effectively improve visitor behaviour to reduce disturbance (Allbrook and Quinn, 2020). One study investigated the effectiveness of using wardening in addition to signage (Medeiros et al., 2007). Wardening is likely to be beneficial for sites experiencing higher visitor numbers and may be relatively resource-efficient if targeted at times of highest visitor traffic (weekends/peak holiday season). For most sites it is likely that using a combination of actions will be most effective.
Regarding inshore waters neighbouring seabird colonies, the studies we reviewed focussed on the behavioural responses of seabirds to approaching vessels, with potential management measures including set-back distances (nearshore exclusion zones) and speed limits. Studies generally found that higher speeds and nearer approaches were associated with larger behavioural response (Ronconi and Clair, 2002; Chatwin et al., 2013; Vogrin, 2013). Coastal buffers with exclusions zones may be possible to implement under existing legislation in Scotland as site-based management measures for SPA seabird colonies. Even relatively small buffers (tens to hundreds of meters) may be able to substantially reduce disturbance. This would require careful site-specific consideration, including inter alia the safety of water users (e.g. potentially increases danger for kayakers and stand-up paddleboarders if required to make wide detours into deeper waters further from the coast), impacts on visitor access (e.g. wildlife watching vessels), and some economic activity (e.g. creel fishing). Most of these impacts can likely be mitigated, for example, through exclusion zones only including the area closest to the coast with a wider zone including some restrictions (e.g. speed limits and craft type).
An increasing potential source of disturbance comes from UAVs (also known as drones), which may be operated by tourist visitors or for research purposes. Only one of the studies included in our review considered these (Bishop et al., 2022), which found contrasting behavioural responses between species, with Common Guillemot not showing any response while Black-legged Kittiwake showed rates of flushing. Counter-intuitively that study found a smaller UAV led to stronger reactions than a larger platform UAV. There are various regulations and recommendations for UAV use (Edney et al., 2023).
While several of the studies reviewed found species-specific responses, it may not be optimal to tailor management and regulation to specific species. Chatwin et al. (2013) suggest that a standard regulation (e.g. a single set distance for all species and all vessels) is easier for visitors to follow and more likely to result in a positive response.
As has been outlined, most studies of disturbance have relied on monitoring behavioural reactions, however, how these behavioural responses relate to population level impacts is generally unclear (Blanc et al., 2006). However, understanding how these sub-lethal effects lead to population level responses can be investigated via modelling studies, as has been used to investigate the impacts of displacement and barrier effects from offshore wind farms (Searle et al., 2018). Even if no behavioural reaction is observed there is still potential for sub-lethal effects, a study on Humboldt Penguin (Spheniscus humboldti) found elevated heartrates when people approached individuals (Ellenberg et al., 2006). This suggests that we should not assume that there is no disturbance just because there is no obvious behavioural response. While it is typically more challenging to monitor burrow or cavity nesting species, it is important to consider these species as they can be vulnerable to visitor disturbance (Bancroft, 2009; Watson et al., 2014; Watson et al., 2021).
It should also be noted that while in general we expect visitors to have some level of negative impacts on seabird populations, in some cases there may be beneficial impacts too. A study on Common Guillemot in Sweden found that removing visitors during the Covid-19 pandemic led to increased predation and disturbance by White-tailed Eagle (Hentati-Sundberg et al., 2021a). Such counter-intuitive effects, mean that any proposed measures should be considered carefully with a site and species-specific context considered.
5.6.6 Knowledge gaps and future directions
Overall, there is a lack of evidence of how behavioural changes impact demographic parameters. There is a need to better understand sub-lethal effects in seabirds in general, which is particularly relevant for understanding disturbance impacts. As has been noted above this can be investigated using bio-energetic models and our physiological understanding can be improved by use of dataloggers that record physiological metrics, like heartrate, or monitoring bio-markers of stress (e.g. corticosterone, see Watson et al. (2021)).
The majority of studies found here were from outwith Scotland and the UK, meaning that there is increased uncertainty when transferring these results to a Scottish context. There would be benefit in more studies on seabirds in the UK, especially on gulls and Razorbill which we found fewest studies for amongst the focal species.
5.6.7 Conclusion
The studies reviewed show that disturbance impacts at seabird colonies are highly species- and site-specific. Overall, there is relatively little specific relevant evidence available to appraise the potential benefit in conservation actions to reduce disturbance, though of those studies reviewed most did find a beneficial effect. Disturbance is hard to measure and relate to population level impacts, especially as responses are not always visible (e.g. stress), and behavioural studies are difficult to translate into effects on demography to understand population level impacts. This review suggests that there could be benefits to actions to reduce disturbance at seabird colonies. However, given the uncertainty in the population level benefits from these, and the species-species aspects, it would require further work to both understand impacts and what mitigations are most effective to have potential for use as strategic compensation. Given that disturbance is a highly site-specific, it would likely have most benefit investigated as project-specific compensation underpinned by detailed knowledge of target sites.
5.7 Ecological feasibility: Reduction of disturbance (at sea)
5.7.1 Summary
The systematic literature review and respective analyses provide evidence that, with a medium degree of confidence, the management of at-sea disturbance has a low to medium effect on at least one demographic parameter of seabirds. However, these findings should be interpreted carefully, as the results from these studies primarily focussed on behavioural aspects, making it challenging to translate such effects to impacts on demographic parameters (e.g. productivity and survival). As such, at this stage, we advise that there is not sufficient evidence to justify its progression as a potential strategic compensatory measure.
5.7.2 Introduction and background
Seabirds have evolved to spend most of their lives in marine environments (Young and VanderWerf, 2023). Throughout the breeding season, seabirds congregate at colonies on land but continue to rely primarily on marine resources for sustenance. Outside of the breeding season, they disperse widely, often covering extensive distances and having prolonged periods at sea without returning to land. The non-breeding season is crucial for resting, moulting, migrating and replenishing energy prior to the breeding season. Consequently, understanding and assessing the threats faced by seabirds during their time at sea is crucial for their effective conservation (Lieske et al., 2020).
Some of the main threats that seabirds encounter whilst at sea include bycatch, overfishing, winter storms, pollution, and disturbance (Yorio et al., 2010; Dias et al., 2019; Lieske et al., 2020). Following Coetzee and Chown (2016) and Allbrook and Quinn (2020), human disturbance to wildlife refers to any activity, event, or action conducted by humans that induce physical, behavioural, or physiological changes in individuals, potentially leading to short- or long-term stress or fitness responses. The primary source of potential disturbance to seabirds at sea is via vessel-related disturbance, affecting seabirds during various stages of their annual cycle, both during the breeding and non-breeding seasons (Bellefleur et al., 2009; Burger et al., 2019).
The presence and activities of different types of vessels, such as fishing vessels, commercial ships, cruise ships, and recreational boats, can disrupt the behaviour and foraging activities of seabirds (Velando and Munilla, 2011; Lieske et al., 2019). Noise, light and oil pollution, physical disturbance caused by vessel movements, and direct collisions can adversely affect an individual’s foraging efficiency, especially during peak chick-rearing period, which may result in increased energy expenditure (Cianchetti-Benedetti et al., 2018; Lieske et al., 2020). Furthermore, vessels can have wider ecosystem effects (Abdulla and Linden, 2008), including displacing seabirds from important foraging locations, alter migration patterns and degrade habitats, potentially disrupting different aspects of the food chain, including their prey. Collectively, these impacts may lead to overall reduced reproductive success and survival among individuals.
Studying seabirds at sea is fundamentally challenging, due to the remote areas and difficult conditions, thus requiring specialised approaches including expensive research vessel access (Ballance, 2007). In addition to this disturbance can be challenging to detect as if there is no apparent behavioural response there can still be physiological impacts such increased stress and cardiac rhythm, and reduced immunity (Ellenberg et al., 2006). This has limited our understanding of how vessels at sea may disturb seabird populations. It is important to understand this risk though, given that vessel activity is increasing, with potential for increased risk to seabirds. Therefore, earlier studies have often focussed on expert judgement and vulnerability assessment approaches to quantify vessel disturbance risk (Certain et al., 2015; Lieske et al., 2019). However, with improving technology and remote monitoring it is becoming more possible to understand how birds respond to vessel presence at larger scales (e.g. Burger et al. (2019)). Certain management options are possible to mitigate for vessels disturbance which could reduce impacts on seabird populations.
The aim of this literature review is to gather evidence on the potential for at-sea vessel management to reduce disturbance impacts on seabirds and to what extent this could lead to population level benefits.
5.7.3 Methods
The conservation action, the reduction of disturbance at sea, is similar to the previous conservation action (see Ecological feasibility: Reduction of disturbance (at colony)). This review, however, will focus on the period when individuals are at sea, away from the colony during both the breeding and non-breeding period. It mainly focusses on marine vessels and on what management actions can be undertaken to decrease their impact on seabirds at sea. Therefore, the review was associated with two research questions. The first research question, ‘What are the potential population level benefits from at-sea vessel management?’, investigates the impact of disturbance management actions on seabirds whilst at sea. The second question, ‘Which management action on what type of vessel provides the strongest benefit?’ aims to identify the management action, as well as the type of vessel, that has the strongest beneficial effect on seabirds.
During a preliminary search, during the refinement of the search strings, it was clear that there was going to be limited evidence on the topic, as most studies focussed on the impact of at-sea disturbance on seabirds, rather than the effect of a specific management action. This observation is consistent with the findings of Sutherland et al. (2021), which also observed a lack of evidence regarding a list of management actions relating to at-sea disturbance on birds. Therefore, we focussed the search on two management actions: reducing vessel speed limits and shipping lanes.
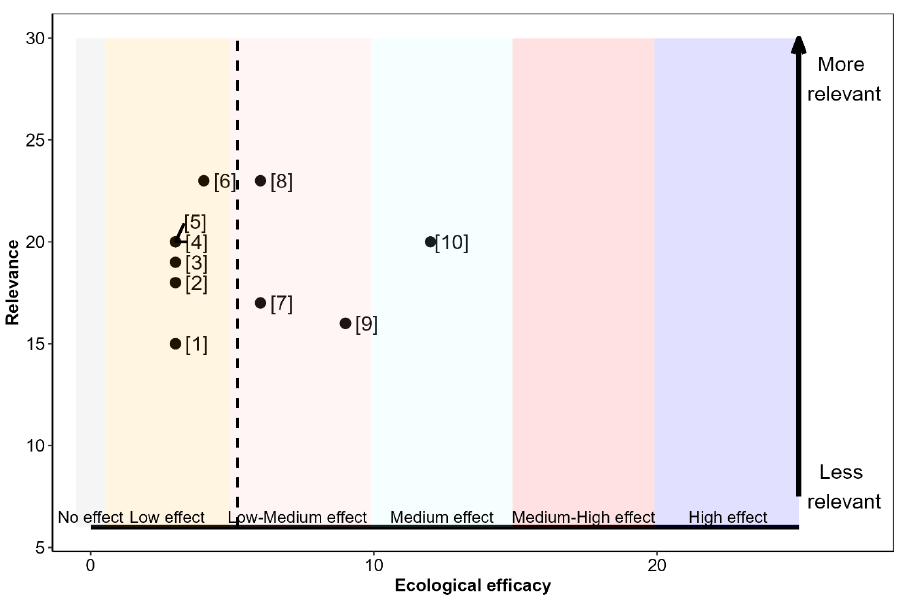
A detailed description of the research question, keywords, search strings, and study selection (Figure 25) is provided in section 8 of Annex 1. Selection criteria can be found in Table A 2.
Information on the time and date that search strings were used, as well as the number of records retained for screening can be found in Annex 2.
Information on the complete reference list obtained during the literature search, and the level at which each reference was eliminated can be found in Annex 3.
Overall, 11 references were included for the final review.
5.7.4 Results
5.7.4.1 General observations
We identified 11 references that met our criteria of measuring the effect of at-sea disturbance on seabirds and waterbirds, all published in English. The literature review yielded relatively current publications ranging from 2005 to 2021 (Figure 26A), with a median publication date of 2011. All records were research articles. Studies mostly conducted behavioural observations and counts from vessels, airplanes, or in land to directly test the degree of impact of the management either on seabirds and/or waterbirds. The assessed studies focussed on the impact of at-sea disturbance on behaviour responses (e.g. whether they flew, dove, no response, distance moved, flush rates; 82% of studies), abundance and density (27% of studies), distribution (27%), foraging activities (18%), and resettlement rates (18%) (Figure 26B). It is important to note that all these are not demographic parameters per se, but potential drivers of demographic parameters and, therefore it is difficult to understand the impact that behavioural changes at sea will have on demographic parameters that will impact populations directly, such as productivity and survival.
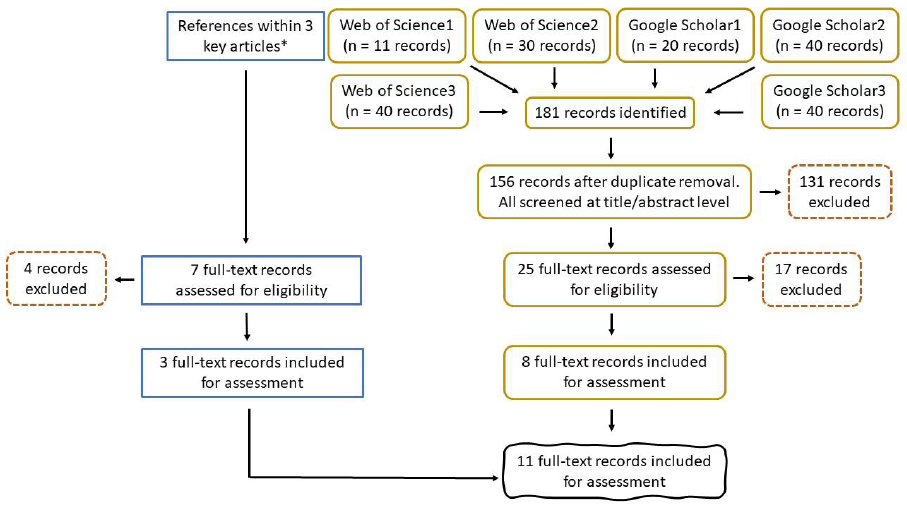
Most studies were undertaken during the non-breeding season (Figure 26C). Studies researched the effect of at-sea disturbance on diverse seabird and waterbird species groups (Figure 26D); mostly on waterbirds (e.g. grebes, ducks, scoters, divers), followed by auks (e.g. Guillemot, Razorbill, Black Guillemot, Marbled Murrelet, Kittlitz’s Murrelet) and cormorants (e.g. European Shag, Rock Shag, Imperial Cormorant), followed by gulls (e.g. Kittiwake), terns (e.g. Common Tern, Arctic Tern), and other species like Fulmar and Gannet . Note that most records researched multiple seabird and/or waterbird species.
Records explored a range of management types (Figure 26E). Specifically, nine studies explored the effect of vessel traffic on birds, three studies explored the degree of vessel disturbance at different speeds, one study focussed on the effect of shipping lanes, and another investigated the extent of vessel disturbance at different distances. Due to the nature of this conservation action, these management types were all indirectly tested. For instance, studies opportunistically observed vessels operating at different speeds and subsequently measured their impact, albeit without directly testing the management type (e.g. ‘reducing speed limits’). Data was collected from multiple seas, coasts, channels, and bays across the globe, close to Germany, Spain, Greenland, Ireland, the UK, Denmark, Argentina, Canada and the USA (Figure 26F). Only one study was undertaken within the UK, specifically in Orkney and investigated the behavioural responses of waterbirds to marine traffic.
5.7.4.2 Evidence scoring
The relevance, as well as the ecological efficacy scores for all 11 references are shown in Table 38 (refer to Annex 4 for a detailed breakdown of the extracted data for each study). Overall, references were of low-medium relevancy for our assessment, with only one study surpassing 20 points. Scores ranged between 12 and 23 out of a maximum possible score of 30, with a median score of 14 and a mean score of 16. For this reason, studies varied in their associated weight, ranging from 0.070 to 0.134 (Table 38). Out of the assessed studies, all but two suggested that vessel management to reduce disturbance could, potentially, have a positive effect on at least one demographic parameter of seabirds or waterbirds.
Table 38. Relevance and ecological efficacy scores, as well as the management action and its effect on seabirds or waterbirds for each assessed study of the disturbance management at sea review. Relevance scores (in bold) could have a potential score between 6 and 30 (6=studies of low relevancy, 30=studies of high relevance). Ecological efficacy scores (in bold) could have a potential score between 0 and 25 (lower scores reflect studies that did not find a significant effect of at-sea disturbance management, or those with low statistical inference; higher scores reflect studies that found a strong beneficial effect of at-sea disturbance management with high statistical inference). SI = Statistical inference and DE = Degree of effect. References are ordered from lowest to highest final ecological efficacy scores (total ecological efficacy score x weight).
This table can be viewed in Excel format.
The ecological efficacy, i.e. the likelihood that the management of disturbance at sea is beneficial on at least one seabird or waterbird demographic parameter, varied across studies. Overall, ecological efficacy scores were low, ranging from 0 (the management of disturbance at sea would not have an effect on birds) to 10 (the management of disturbance at sea has a medium effect on bird colonies) out of a maximum possible score of 25. The median score was 6 indicating that the effect varied according to each study (Figure 27). The lower scores observed can be attributed to the fact that the studies primarily focussed on measuring behavioural aspects rather than directly measuring the effects on demographic parameters (e.g. it is unclear how and to what degree higher flushing rates during the non-breeding season affect survival).
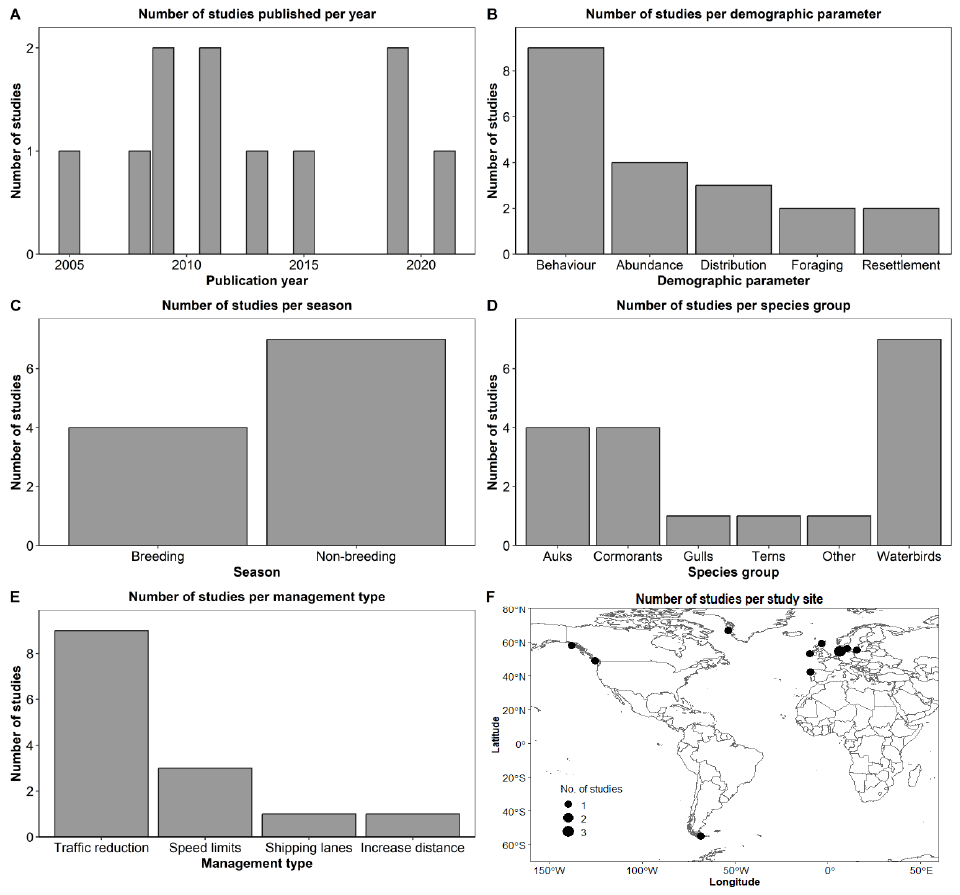
The sum of all final weighted scores is 5.7 (i.e. the mean of ecological efficacy weighted by relevance), indicating that overall, the evidence suggests that at-sea disturbance management will benefit seabirds to a low-medium degree (Figure 27), but this is through indirect evidence and difficult to be certain of – treat with caution.
5.7.4.3 Overall confidence scoring
We estimated the median of the relevance index score, the independence and quantity of the evidence and the concordance between studies to obtain an overall score to assess the confidence in our findings (see Confidence). The total score for this measure was 8 out of the possible 15 (Table 39), suggesting that we have a medium degree of confidence that the assessed estimation of the ecological efficiency is an accurate reflection of the true ecological efficacy of the management of at-sea disturbance.
Table 39. Assessment of the confidence in the evidence for management of at-sea disturbance.
This table can be viewed in Excel format.
5.7.4.4 Main findings: the effect of avian predator management
In summary, our literature review and respective analyses provide evidence that, with a medium degree of confidence, the management of at-sea disturbance has a low-medium effect on at least one demographic parameter of seabirds. This, however, needs to be treated with caution as the results from these studies are mainly behavioural, making it challenging to accurately quantify their impacts on demographic parameters (e.g. productivity and survival).
The key findings from the most relevant studies are summarised in Table 40. Most studies indirectly tested the effect of vessel management at sea on seabirds and/or waterbirds. While these studies did not implement explicit management actions (e.g. setting speed limits), they compared the responses of birds under different conditions (e.g. response to boats that were going at different speeds). Results from these studies can therefore provide insights into the potential effects of specific management actions.
Most studies found that birds were more abundant with no or little concurrent ship traffic, this is proven with Red-throated divers (Burger et al., 2019), European Shags (Velando and Munilla, 2011), Marbled Murrelets (Bellefleur et al., 2009) and other waterbirds (Schwemmer et al., 2011). Red-throated divers flushed shorter distances when fewer ships were present (Burger et al., 2019). European Shags, Common Eiders and Kittlitz’s Murrelets reduced foraging once a boat was within the vicinity and, in most cases, birds flew away (Agness et al., 2008; Merkel et al., 2009; Velando and Munilla, 2011). These events increased with higher boat traffic, substantially reducing the time spent foraging. This also meant that when more boats were present, the probability of individuals being excluded form the best feeding areas also increased (Velando and Munilla, 2011). In the case of Common Eiders, there was a cumulative effect of repeated disturbances on the feeding activity if disturbances were close in time and space (Merkel et al., 2009). Kittlitz’s Murrelets spent more time loafing and less time diving in days when there was no vessel traffic (Agness et al., 2008) and Great Northern Divers didn’t seem to flush in response to boat
Table 40. Summary findings of the five most relevant studies on the effect of the management of at-sea disturbance on seabirds and waterbirds, in order of decreasing relevance.
This table can be viewed in Excel format.
traffic, even when these passed within <20 m from individuals (Gittings et al., 2015). Faster and closer boats caused a greater proportion of birds to flush, and at further distances on (Agness et al., 2008; Bellefleur et al., 2009; Larsen and Laubek, 2013; Burger et al., 2019).
Schwemmer et al. (2011) studied the impacts of shipping lanes on waterbird species in German offshore waters and found that divers preferred areas located between shipping lanes while actively avoiding the shipping lanes themselves. Some indications of habituation were observed within the shipping lanes, as certain birds did not flush in response to passing ships. However, it remains uncertain whether waterbirds can fully habituate to the presence of freely moving ships.
Fliessbach et al. (2019), however, found that their estimated Ship Traffic Disturbance Vulnerability Index varied strongly amongst species with Black Guillemot being more susceptible to vessel disturbance followed by Razorbill, Guillemot, Gannet, Kittiwake, gulls, and terns. Similarly, Jarrett et al. (2021) found differences in the likeliness and behaviour to respond to passing vessels; Red-throated Diver, Black-throated Diver, and Slavonian Grebe, for example, were most likely to respond compared to other waterbirds, and while Red-throated Diver and Slavonian Grebe were highly likely to flush, Black-throated Diver and Great Northern Diver were more likely to swim or dive.
5.7.4.5 Biases or Conflict of interests
Of the 11 studies, three include potentials for conflicts of interests (note we do not make judgement on whether these lead to an actual conflict of interest or influence study findings but provide the information for the readers’ awareness). Two studies were wholly or partly funded by Federal Ministries (Schwemmer et al., 2011; Burger et al., 2019) and one article was funded by a Harbour Company (Gittings et al., 2015). The authors of the other studies did not disclose any potential biases or conflicts of interest.
5.7.5 Discussion
We did find evidence that seabirds could benefit from reducing at-sea disturbance. However, this was based on relatively few studies (only 11 met our study inclusion criteria), and the majority of these looked at behavioural responses rather than measures that are closer to population level impacts. This is akin to the findings we encountered during the Ecological feasibility: Reduction of disturbance (at colony) review, where there were the same general limitations with the evidence base available.
For this study, we focussed on the focal species and similar seabird species, however, given the lack of studies, we also included waterbird species such as Diver species (Gavia spp.) and sea ducks for which there was more evidence available, see e.g. (Merkel et al., 2009; Schwemmer et al., 2011; Burger et al., 2019). Diver species are known to be particularly vulnerable to vessel activity showing strong displacement responses, so have been the focus of various studies around their behavioural responses and their energetics (to better understand potential for physiological impacts) (Burger et al., 2019; Vilela et al., 2021; Thompson et al., 2023). However, whether this leads to a population level impact is unclear (Vilela et al., 2021). When considering management measures for seabirds aiming to reduce at-sea disturbance, it is important to also take into account these other species groups. This involves ensuring that any measures implemented do not lead to unintended consequences for other species, but also to learn from measures developed for better-studied groups.
There are a range of potential vessel management measures that could reduce disturbance. Bellefleur et al. (2009) identified three broad potential actions (in the context of reducing disturbance on Marbled Murrelets) most of which have also been considered in other studies (Lynch et al., 2010; Burger et al., 2019; Studwell et al., 2021): (1) limiting boat speed in nearshore areas; (2) spatial management of vessels to direct activity away from areas with the highest bird densities (e.g. using shipping lanes or area closures); (3) seasonal closures to boat traffic during periods of greatest activity. In addition to these broad measures that could apply to all vessels, large infrastructure projects, such as offshore wind farm construction and maintenance are often required to develop vessel management plans to mitigate the risk of disturbance.
The regulation and management of specific aspects of vessel traffic have the potential to provide some benefits in terms of survival and productivity in seabirds but there is a lack of specific evidence, especially to be able to predict the level of benefit to specific species. Evidence suggests that some species are more vulnerable to vessel activity than others (Fliessbach et al., 2019; Jarrett et al., 2021), though this is based on behavioural responses, so whether these translate to greater population level impacts is unclear. As discussed in further detail in Ecological feasibility: Reduction of disturbance (at colony), it is challenging to directly link the effects of disturbance and decreased feeding to population level responses. Studies using novel biologging devices to record behaviour and physiology at sea (Ropert-Coudert et al., 2012) can provide more direct measures than observational studies, e.g. of heart rate which is related to stress responses and energy expenditure (e.g. Ellenberg et al. (2006)). Behaviour and energetics of free-moving seabirds could then be related to vessel presence and movements, in a similar way to what has been done for studies relating to interactions with fishery vessels (Votier et al., 2010). This general approach has been applied for Red-throated Diver in Burger et al. (2019). There is potential to apply this to a wider suite of species, and, depending on the tracking already collected, it may be possible to do such analyses using existing datasets.
5.7.6 Conclusion
Overall, while our review indicates that reducing disturbance at sea would benefit seabirds, there was insufficient evidence to be able to come to clear conclusions on what actions may be helpful and what level of benefit could be anticipated. There is evidence of vessel presence and activity leading to behavioural response in marine birds, but few studies have gone on to link this to population level impacts. As such, at this stage, we advise that there is not sufficient evidence to be able to progress as a potential strategic compensatory measure. However, given that there is evidence that vessels can affect seabird behaviour, we advise that further research should be conducted to understand how disturbance effect varies between species, whether this leads to population level responses, and what actions may best reduce impacts.
Contact
Email: ScotMER@gov.scot
There is a problem
Thanks for your feedback