Sandeel fishing consultation: review of scientific evidence
The review of scientific evidence that has been used to inform the development of proposals to close fishing for sandeel in all Scottish waters.
4 Seabirds
Scotland holds internationally important numbers of breeding seabirds, with 24 species regularly breeding in Scotland (Mitchell et al. 2004). A large proportion of these species include sandeel in their diet during the breeding season. The ability of seabirds to prey on sandeel will depend on both the absolute numbers of sandeel (stock biomass) and the availability of sandeel to seabirds. Seabirds are constrained in both the distance from nest sites that they can forage (when breeding) and the depth in the water column that they can reach, with both foraging range and dive depth varying greatly among species.
4.1 Sandeel stock biomass and availability to seabirds
The estimated sandeel total stock biomass (TSB) in the SA4 region off eastern Scotland, has fluctuated greatly over the last seven decades (ICES 2022, Figure 21). Drivers of sandeel stock biomass are discussed in detail in section 2 and include bottom-up processes such as environmental effects on sandeel copepod prey and density dependent regulation, and top-down effects such as natural predation by piscivorous fish, marine mammals and seabirds, as well as fishing.
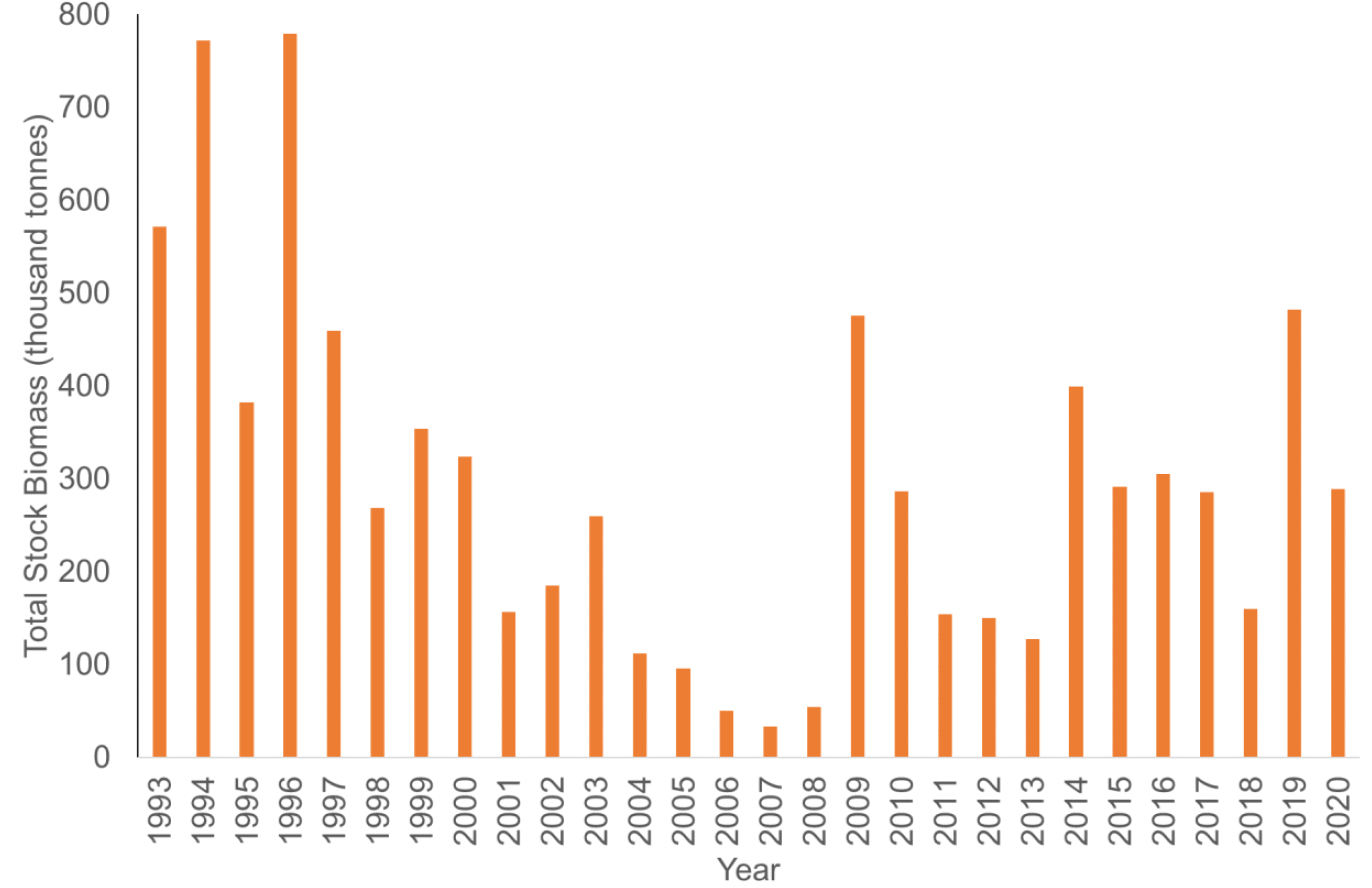
4.1.1 Sandeel in the water column and availability to different seabird species
Following the planktonic larval stage, sandeel are only in the water column for a few months of the year. The extent to which seabirds can dive down into the water column to obtain sandeel at different depths varies greatly among species. Surface feeding seabirds, such as terns (Sterna sp) and black-legged kittiwake (Rissa tridactyla, hereafter kittiwake), can only take fish very close to the surface whereas other species such as common guillemot (Uria aalge, hereafter guillemot), razorbill (Alca torda) and Atlantic puffin (Fratercula arctica, hereafter puffin) can dive to considerable depths. Guillemot and European shag (Gulosus aristotelis, hereafter shag) can also extract fish from the sediment on the seafloor and so can feed on sandeel even when they are not in the water column.
The period during which sandeel are in the water column shows inter annual variation. The timing of the sandeel' seasonal return to the sediment is highly important for surface feeding seabirds as an early return to the sediment can result in sandeel no longer being available towards the end of the seabird breeding season.
4.2 Prevalence of Sandeel in Seabird Diet
The extent to which seabirds are dependent on sandeel varies among species. For example, guillemot have been shown to have greater capability to switch to foraging on sprat and small herring when sandeel are unavailable compared to kittiwake (Rindorf et al. 2000). See Table 2 for a list of seabird species known to prey on sandeel.
During breeding many Scottish seabird populations exploit seasonal peaks in sandeel abundance, feeding on both adult (1+ year group) and juvenile (young-of-the-year; age 0) age classes. For example, on the Isle of May, Firth of Forth, which is one of the best studied seabird colonies in the world, sandeel comprised approximately 75% of the breeding season diet of shag, kittiwake and puffin between 1991 and 2011 (Newell et al. 2013). Sandeel also constituted a substantial proportion of the diet of seabirds breeding on Canna, Western Isles between 1981–2007 (Swann et al. 2008). Over this period sandeel were the greatest component (62%) of regurgitations from young shag and adults feeding chicks, comprised a quarter of young guillemot diet, and occurred in 60% of kittiwake regurgitations, being the commonest fish family in 14 of the 20 years in this latter seabird species' diet (Swann et al. 2008).
Many seabirds in Scotland display seasonal shifts in diet composition from larger adult to smaller juvenile sandeel throughout breeding (Phillips et al. 1999; Lewis et al. 2001; Lewis et al. 2003; Wilson et al. 2004; Howells et al. 2017; Harris et al. 2022), which is consistent with sandeel life history (Winsdale 1974). For example, adult kittiwake generally switch from feeding on older sandeel at the start of breeding (April/May) to juvenile sandeel (for both self-feeding and their young) as the season progresses (Harris & Wanless 1997; Lewis et al. 2001). Similarly, in shag the proportion of adult sandeel decreases and juvenile sandeel increases throughout the breeding season (Howells et al. 2017). Spatial variation in diet composition is also apparent, with the importance of sandeel varying among seabird colonies of the same species, likely linked to local environmental conditions, foraging distributions and prey availability (Bull et al. 2004; Anderson et al. 2014; Wilson et al. 2021).
Due to the challenges of studying seabird diet in the non-breeding season, when birds are at sea and away from their colonies, less is known about the importance of sandeel in seabird diet during this period (Barret et al. 2007). Despite this, the limited information that exists suggests that seabirds forage on a wider variety of prey during the non-breeding season, but still include sandeel to some extent (Blake 1985; Howells et al. 2018). For example, Howells et al. (2018) identified that the frequency of sandeel occurrence in shag diet decreases markedly between the breeding and non-breeding season. Using stomach contents from shot birds, Blake (1985) also identified differences between the breeding and non-breeding seasons, with sandeel dominating between March and August and a changeover occurring in September when clupeid and gadoid remains became increasingly frequent. Thus, while sandeel are preyed upon by Scottish seabirds throughout the year, other prey are also important in the non-breeding season to a varying extent.
While sandeel have traditionally been considered one of the most abundant and energy rich prey for seabirds in Scotland (Dunnet et al., 1990; Hislop, Harris and Smith, 1991; Furness and Tasker, 2000), the availability, size and calorific content of this species has declined in recent decades (Wanless et al., 2004, 2005; Frederiksen et al. 2013; Wanless et al. 2018). As a result, many seabird populations now appear to have a reduced dependency on sandeel, although this can be highly variable among years and colonies (Wanless et al. 2004, 2005; Heubeck 2009, Anderson 2018; Howells et al. 20017, 2018; Wanless et al. 2018). For example, in a comprehensive long-term, community-level analysis of seabird diet on the Isle of May between 1973 and 2015, Wanless et al. (2018) identified reductions in the importance of sandeel in seabird diet, along with a decline in adult (age 1+) relative to juvenile (age 0) sandeel. Further, by combining data on sandeel length and energy density in the diet of seabirds, Wanless et al. (2018) identified overall reductions in average energy values of both adult (c.70% reduction) and juvenile (c.40% reduction) sandeel over the past 4 decades. This reduction was detected in both inshore and offshore foraging seabird species, suggesting a widespread pattern of change. While concurrent, independent time series data on prey abundance at a spatial scale relevant to seabird breeding foraging ranges are lacking, the available fisheries data, collected over a much wider area and coarser resolution, were broadly in line with these observed changes - indicating that sandeel have become significantly shorter in length and lighter than those in central and north-eastern areas (Rindorf et al. 2016; Wanless et al. 2018). Whilst energy density (kJg-1) per gram of sandeel did not decline between 1973-2015 (Wanless et al. 2018), the absolute amount of energy per fish will be lower in smaller fish than larger fish (Booth et al. 2023, Wanless et al. 2004), meaning seabirds need to catch more, smaller sandeel to obtain the same energy to meet their own and their offspring's energetic requirements (Rindorf et al. 2016). Where such compensation is not possible, seabird chicks of many species, including guillemot, shag, kittiwake, puffin and razorbill, experience net reductions in energy value at the time of chick-rearing because of declines in sandeel length (Burthe et al. 2012). However, some species may be able to maintain their energetic intake for chicks through prey switching (Smout et al. 2013). Indeed, in Shetland, declines in local sandeel biomass resulted in great skua switching from feeding predominantly on sandeel via kleptoparisitism of other seabird species to predating other seabirds (Votier et al. 2007), highlighting the complexities of marine food webs and the challenges of predicting the response of seabirds to changes in sandeel populations.
4.3 Predicted vulnerability of seabirds to declines in sandeel abundance
Due to differences in seabird life-history, ecology and diet, the dependency on and vulnerability to changes in sandeel biomass and availability varies among species (Rindorf et al. 2000; Furness & Tasker 2000). Furness & Tasker (2000) assessed the vulnerability of the North Sea seabird community to reductions in sandeel abundance using an 'index of vulnerability to reduced food-fish abundance' based on a suite of species characteristics including: body size, foraging costs and range, diving ability, spare time in daily energy budget and dietary flexibility. Some species, such kittiwake and Arctic skua, are considered highly sensitive to changes in sandeel availability due to factors such as high foraging costs, limited diving ability (i.e., surface feeders) and restricted dietary flexibility. Other species, such as shag and gannet, are considered more resilient to fluctuations in sandeel abundance, due to greater foraging and dietary flexibility, with gannet far ranging and shag able to target prey throughout the water column (Table 2).
| Species | Small Size | High cost of foraging per unit of time | Constrained to short foraging range | Little ability to dive | Lack of spare time in daily budget | Low ability to switch diet | Score |
|---|---|---|---|---|---|---|---|
| Arctic tern | 4 | 4 | 4 | 3 | 4 | 3 | 22 |
| Roseate tern | 4 | 4 | 4 | 3 | 4 | 3 | 22 |
| Little tern | 4 | 4 | 4 | 3 | 4 | 2 | 21 |
| Common tern | 3 | 4 | 4 | 3 | 4 | 2 | 20 |
| Sandwich tern | 3 | 4 | 3 | 3 | 3 | 3 | 19 |
| Black-legged kittiwake | 2 | 2 | 1 | 4 | 4 | 3 | 16 |
| Arctic skua | 2 | 2 | 3 | 4 | 1 | 3 | 15 |
| Black-headed gull | 3 | 3 | 3 | 4 | 2 | 0 | 15 |
| Common gull | 2 | 3 | 3 | 4 | 2 | 0 | 14 |
| Black guillemot | 2 | 3 | 3 | 1 | 3 | 2 | 14 |
| Great skua | 0 | 3 | 3 | 4 | 1 | 2 | 13 |
| Atlantic puffin | 2 | 3 | 1 | 2 | 3 | 2 | 13 |
| Razorbill | 1 | 3 | 2 | 1 | 2 | 3 | 12 |
| Red-throated diver | 0 | 3 | 4 | 0 | 2 | 3 | 12 |
| Lesser black-backed gull | 1 | 2 | 2 | 4 | 1 | 1 | 11 |
| Herring gull | 1 | 2 | 3 | 4 | 1 | 0 | 11 |
| Greater black-backed gull | 0 | 2 | 3 | 4 | 1 | 0 | 10 |
| British storm petrel | 4 | 2 | 1 | 3 | 0 | 0 | 10 |
| Leach's petrel | 4 | 2 | 1 | 3 | 0 | 0 | 10 |
| Common guillemot | 1 | 3 | 1 | 0 | 2 | 2 | 9 |
| Shag | 0 | 3 | 3 | 0 | 0 | 2 | 8 |
| Great cormorant | 0 | 3 | 4 | 0 | 0 | 0 | 7 |
| Manx shearwater | 2 | 1 | 0 | 2 | 0 | 2 | 7 |
| Northern fulmar | 1 | 0 | 0 | 4 | 2 | 0 | 7 |
| Northern gannet | 0 | 2 | 0 | 2 | 1 | 0 | 5 |
| Species | Vulnerability Score | Proportion of sandeels in diet | Sensitivity to sandeel abundance |
|---|---|---|---|
| Kittiwake | 16 | 0.87 | 13.9 |
| Puffin | 13 | 0.81 | 10.5 |
| Razorbill | 12 | 0.77 | 9.2 |
| Shag | 8 | 0.96 | 7.7 |
| Guillemot | 9 | 0.84 | 7.6 |
| Tern species | 21.8 | 0.34 | 7.4 |
| Gannet | 5 | 0.42 | 2.1 |
Daunt et al. (2008) further added to Furness & Tasker's (2000) vulnerability index by using data on the proportion of sandeel in the diet of species breeding at the Isle of May colony during 1996-2003, to assess the sensitivity of species to sandeel abundance (Table 3). This sensitivity score is a measure of both the seabird species' vulnerability to a paucity of sandeel and their reliance on sandeel.
If the predictions of Furness & Tasker (2000) and Daunt et al. (2008) are correct, the seabird species most sensitive to changes in sandeel abundance should have exhibited a decrease in breeding success in response to reduced sandeel size and abundance over recent decades. Kittiwake and puffin would be predicted to have shown the greatest decrease in breeding success over recent decades while guillemot and gannet would be predicted to have not shown the same declines.
4.4 Seabird demography and sandeel abundance, biomass and availability
Understanding the extent to which seabird demography (breeding success, survival, and population size) is determined by sandeel abundance, biomass and availability to seabirds, and hence the potential benefits that a sandeel fishery closure might bring for seabirds, is not straightforward. However, the large changes in sandeel biomass and availability over the last few decades, coupled with intensive monitoring of seabird population size, breeding success and diet, particularly at the Isle of May, enables investigation of any correlation between seabird demography and sandeel biomass.
Furness and Tasker (2000) and Daunt et al. (2008) identified kittiwake as one of the species most sensitive to changes in sandeel biomass. Kittiwake breeding abundance in Scotland has undergone a sustained and significant decline (Harris & Wanless, 1997) with the most recent Seabird Monitoring Programme breeding abundance index, for 2019, being 60% below the 1986 baseline (JNCC, 2021).
4.5 Seabird breeding success and sandeel biomass and abundance
A consistent pattern in the way seabird breeding success changes with forage fish abundance has been reported for many seabird-forage fish interactions around the globe. Known as 'one-third for birds', Cury et al (2011) found seabird breeding success to vary little or not at all at intermediate and high forage fish abundance, but once forage fish abundance dropped below a threshold of one-third of maximum biomass, seabird breeding success rapidly declined. This relationship has also been found for seabirds feeding on sandeel, e.g., for breeding success of Arctic skua, great skua and kittiwake on Foula in relation to the Shetland sandeel total stock biomass (Frederiksen et al. 2007; MacArthur Green 2021). A similar relationship has also been found for a proxy of adult survival at the Isle of May for kittiwakes (MacArthur Green, 2021).
Further evidence supporting the role of sandeel abundance in driving kittiwake breeding success at large spatial scales comes from synchronised variation in breeding success among multiple kittiwake colonies. Frederiksen et al. (2005) found that regional variation in prey availability had a stronger effect on kittiwake breeding success than local prey depletion. Olin et al. (2020) found the spatial structure in sandeel populations played a role in driving this synchrony within and among clusters of kittiwake colonies.
As described above (see section Prevalence of Sandeel in Seabird Diet) seabird species which are capable of switching to feed on alternative prey have generally been able to sustain better breeding success than species without the ability to switch. However, in Shetland, alternative prey such as sprat and juvenile herring are scarce (Frederkisen et al. 2007, Hamer et al. 1993; Furness & Tasker 2000). Consequently, when the Shetland sandeel stock collapsed in the 1990s, many seabird species exhibited reduced breeding success e.g., Arctic skua, kittiwake (Davis et al. 2005; Furness 2007; Phillips et al. 1996; Frederkisen et al. 2007) and survival e.g., Arctic skua, great skua, kittiwake (Davis et al. 2005; Oro & Furness 2002; Ratcliffe et al. 2002). Olin et al. (2020) noted the extent to which sandeel populations drive patterns in kittiwake breeding success among colonies is stronger in Shetland and Orkney than further south, due to the absence of alternative prey.
4.6 Influence of sandeel availability on seabird demography
Seabird breeding success is influenced not only by sandeel biomass, abundance, and quality but also by their availability. Assessing sandeel availability to seabirds, especially surface feeders, is difficult as availability varies as sandeel move between the water column and the sediment.
Kittiwakes on the Isle of May had higher breeding success during 1986-1996 when age 0 sandeel made up a higher proportion of chick diet (Harris & Wanless, 1997). During the late 1980s to late 2000s, community level comparisons of seabird breeding on the Isle of May showed a synchronised decline, driven by reduced chick survival rather than failure during egg incubation (Lahoz-Montfort et al. 2013). Concurrent community-level reductions in the importance of sandeel in chick diet suggests dietary changes could have contributed to the synchronised declines (Wanless et al. 2018). Breeding success on the Isle of May in kittiwake, razorbill and guillemot was greater in years when the proportion of sandeel in chick diet was higher (Searle et al. 2023).
Guillemot, kittiwake and shag breeding success was significantly lower when sandeel fisheries catch per unit effort (CPUE) was lower, suggesting more sandeel in the water column can contribute to higher breeding success (Rindorf et al 2000). Whilst this positive relationship was strongest in surface-feeding kittiwake, it was also found for shag and guillemot which are capable of foraging throughout the water column, implying higher foraging efficiency for these two species when sandeel availability in the water column is greater.
As well as sandeel availability in the water column, temporal availability of sandeel also influences seabird breeding success. The peak in sandeel abundance needs to coincide with the seabird breeding season. Breeding success of three seabird species on the Isle of May was greatest when CPUE in June was high and the May/June CPUE ratio (an index of the timing of the onset of sandeel burying behaviour) was low, implying a peak abundance in May is too early to benefit seabird chicks (Rindorf et al 2000).
The extent to which availability of age 0 and age 1+ sandeel influences breeding success varies among seabird species. Kittiwake, guillemot, puffin, and shag breeding success was positively related to sandeel larval biomass in the previous year, implying seabirds were feeding on 1 year old sandeel (Frederiksen et al. 2006). Kittiwake tend to feed on age 1+ sandeel in April and May, shifting to age 0 sandeel in June and July, with highest breeding success occurring when age 0 sandeel appeared early in the season (Lewis et al. 2001).
Thus, not only is total abundance or biomass of sandeel important to seabirds but also the timing of that peak abundance and the extent to which it coincides with key stages of the seabird breeding season (Rindorf et al. 2000).
Kittiwake breeding success was higher when sandeel size was larger (Lewis et al. 2001) but sandeel in seabird diet have been getting smaller over the last 30 years (Wanless et al 2004). Consequently, seabird breeding success has also been impaired by the size and thereby energy value of sandeel as well as their availability.
4.7 Industrial sandeel fisheries and seabird demography
4.7.1 Challenges of establishing a relationship between industrial sandeel fisheries and seabird demography
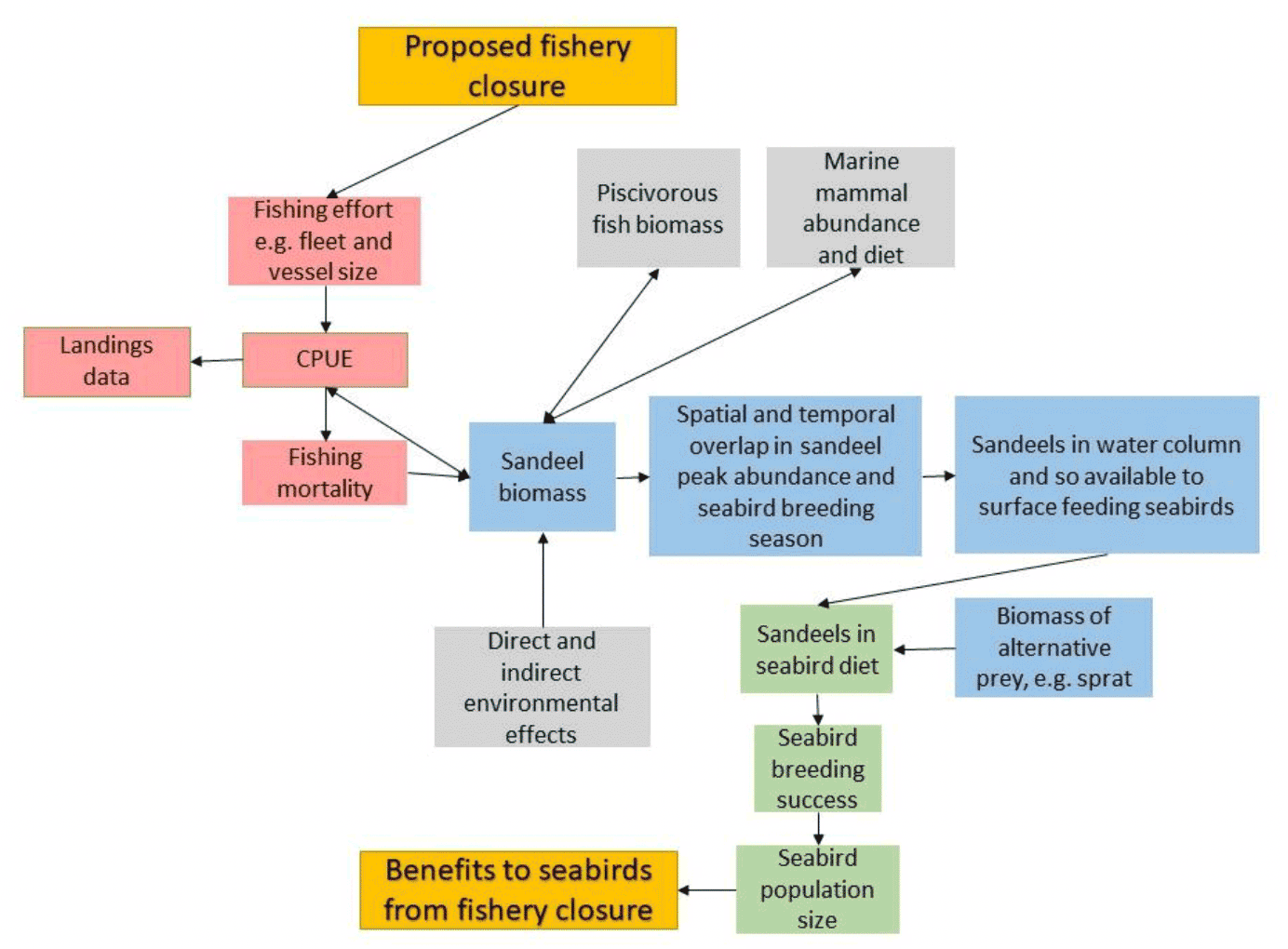
Obtaining measures of how a sandeel fishery changes the abundance or availability of sandeel to seabirds, and hence seabird demography, is not straightforward.Figure 27 illustrates the multiple complexity of linking changes to a sandeel fishery to seabird population size.
Only correlative relationships between breeding success and sandeel fishing and/or abundance can be established, meaning confidence in observed results is less than from an experimental manipulation. Also, seabird demography is driven by lag effects and interannual fluctuations that make teasing out effects of a fishery from environmental variation difficult (Searle et al. 2023).
The sandeel fishery is only one driver of sandeel stock biomass, the latter also being determined by top down (natural predation (Furness 2002)) and bottom up (environment and copepod prey abundance (Frederiksen et al. (2004)) regulation. This makes demonstrating a causal relationship between industrial sandeel fishing and seabird demography challenging (Sydeman et al. 2017).
4.7.2 Seabird demography and sandeel fisheries
Seabird breeding success has been shown to have a non-linear relationship with forage fish biomass, with breeding success rapidly declining as stocks fall below one third of maximum (Cury et al, 2011). Fisheries targeting fish that are key prey for seabirds have the potential to push stocks below this threshold, causing decreases in breeding success (Sydeman et al 2017; Cury et al 2011).
Fishing effort from the Wee Bankie sandeel fishery was associated with a decreased proportion of sandeel in puffin diet (Searle et al. 2023). Fishing effort was also associated with a decrease in the relative proportion of age 0/age 1+ sandeel in diet of kittiwake, razorbill and puffin (Searle et al. 2023). However, only kittiwake breeding success decreased with fishing effort.
Kittiwake breeding success at Flamborough & Filey Coast SPA, eastern England, was positively correlated with sandeel spawning stock biomass the preceding winter and negatively related to fishing mortality with a two-year lag (Carroll et al. 2017). GPS tracking indicated kittiwakes and the fishing fleet were both targeting the same grounds on the Dogger Bank (Carroll et al. 2017).
Whilst seabirds generally feed on age 1+ sandeel during the first part of the breeding season and then switch to age 0 sandeel in May, the fishery targets only age 1+ sandeel in May, June and July. So, whilst there was some spatial overlap in foraging areas used by the fishery and seabirds (the extent of overlap varying among species) (Wanless et al. 1998), they were likely to be targeting different age classes (Daunt et al. 2008).
Fishery impacts on sandeel availability are known to be potentially additive to environmental effects (Daunt et al 2008, Rindorf et al 2000). Cook et al (2014), when investigating the use of seabird breeding success as an environmental indicator, found a negative relationship between kittiwake breeding success and sea surface temperature among colonies along the east coast of the UK. When the Wee Bankie sandeel fishery was active, the number of kittiwake colonies failing to achieve predicted breeding success, given the underlying environmental conditions (e.g., sea temperature), increased, particularly in areas close to the Wee Bankie. Once the fishery closed, colonies failing to achieve expected levels of breeding success instead became clustered around Orkney and Shetland, due to reduced sandeel availability and increased great skua predation (Cook et al 2014). This suggests that fishery impacts on kittiwake breeding success were additive to underlying environmental effects.
4.7.3 Benefit of sandeel fishery closures to seabirds
On two separate occasions, industrial sandeel fisheries have been closed due to concerns about their impacts on the breeding success of seabirds (Greenstreet et al. 2006). During the mid to late 1990s, a small sandeel fishery off Shetland was closed following declines in breeding success of seabirds including Arctic tern (Monaghan et al. 1989), great skua (Hamer et al. 1991) and kittiwake (Hamer et al. 1993).
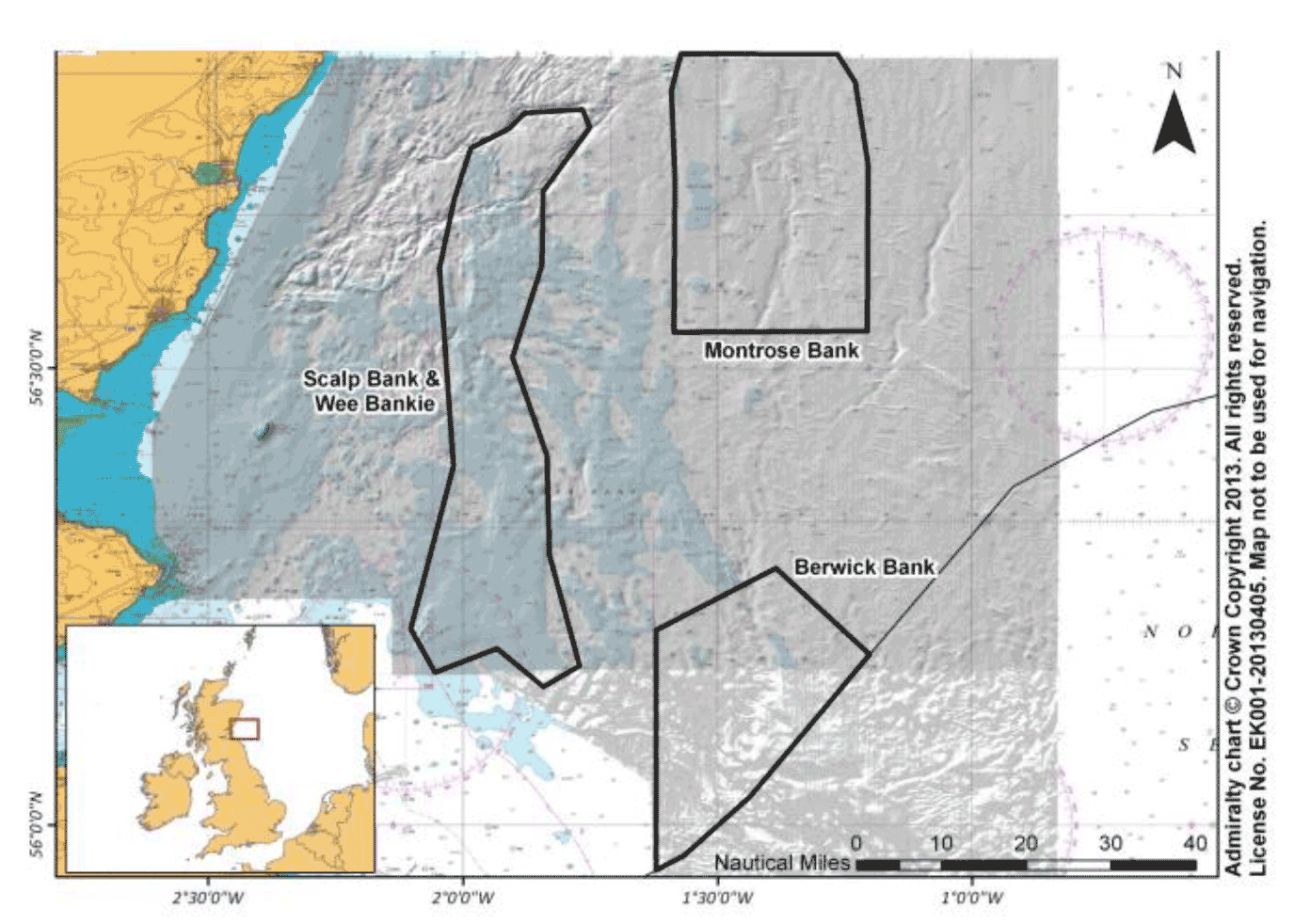
More recently, an industrial sandeel fishery on the Wee Bankie, Scalp Bank and Marr Bank that opened in 1990 was closed in 2000 due to concerns about the fishery impacting breeding success of seabirds nesting around the Firth of Forth, including the Isle of May (Rindorf et al. 2000). Guillemot, razorbill, kittiwake and, to some extent puffin, were known to use the fished area for foraging during the breeding season (Wanless et al. 1998). The evidence for the fishery reducing breeding success, particularly for kittiwake, was so strong that ICES recommended using 'the criterion of kittiwake breeding success falling below 0.5 fledged chicks per well-built nest for three successive seasons as the threshold to close sandeel fisheries in areas important for foraging by the kittiwake colonies being monitored' (ICES, 2000).
4.7.4 Wee Bankie sandeel fishery closure and seabird demography
Following closure of the Wee Bankie sandeel fishery in 2000, sandeel abundance initially increased, as did kittiwake breeding success. Consumption rates of age 0 sandeel were higher after the fishery closure, despite the fishery not targeting age 0 sandeel (Daunt et al. 2008). However, no significant relationship between sandeel abundance and breeding success was found in shag, guillemot, razorbill, puffin, common tern or Arctic tern (Daunt et al. 2008; Frederiksen et al. 2008).
The effect of the opening and subsequent closure of the Wee Bankie fishery on kittiwake breeding success can be investigated using a quasi-experimental approach, with a BACI (before-after-control-impacted-design). The Before/After is the period when the Wee Bankie fishery was active (1991-1998) compared with when the fishery was inactive (1999 onwards); the Control/Impacted uses breeding success at colonies where kittiwakes forage in areas that remain subject to industrial sandeel fishing throughout the study period as a control, and colonies foraging within the closed Wee Bankie fishery zone as the treatment (Frederiksen et al. 2008; Searle et al. 2023). Foraging areas used by birds from impacted colonies overlapped, to varying extents, with the grounds used by the fishery (Wanless et al. 2018; Daunt et al. 2008). The BACI analysis showed that for colonies with kittiwake foraging in the impacted area, breeding success was significantly lower when the fishery was active, compared to when the fishery was inactive, while no significant change in breeding success was found at control colonies over the same time period (Frederiksen et al. 2008). Kittiwake breeding success at impacted colonies was also negatively correlated with fishery effort when the fishery was active but not at control colonies (Frederiksen et al. 2008).
Recently, Searle et al. (2023) repeated the analytical design of Frederiksen et al. (2008), examining changes in breeding success and diet of kittiwake, guillemot, razorbill and puffin during 1986-2018 in relation to the opening and closure of the Wee Bankie fishery. All four species showed a decrease in breeding success across the 32 years of the study. Kittiwake showed a decrease in breeding success attributable to the sandeel fishery (from 52% to 23% proportion of nests to young fledged), followed by a smaller increase (10%) when the fishery closed, compared with control colonies . Importantly, closing the fishery did not enable kittiwake breeding success to recover to pre-fishery levels. Puffin, razorbill and guillemot showed no negative effects of the fishery on breeding success, nor any positive effects following closure (Searle et al. 2023).
4.8 Other drivers of seabird demography
Evidence of sandeel biomass, abundance and availability influencing seabird breeding success, and how this changed in the presence and absence of the Wee Bankie sandeel fishery, is presented above. However, seabird breeding success and population size are also driven by other factors. There are numerous other pressures on Scottish seabird populations including Highly Pathogenic Avian Influenza (NatureScot 2022) and climate change (Mitchell et al. 2002). Wind and rain can reduce breeding success in kittiwake (e.g. Christensen-Dalsgaard et al. 2018; Alvestad 2015; Newell et all 2015) although changing environmental conditions are more likely to influence prey quality and availability rather than have substantial direct impacts on seabirds (MacDonald et al. 2015). Despite synchrony in kittiwake breeding success (Olin et al. 2020; Frederiksen et al. 2005), breeding success can vary between colonies in close proximity due to other effects such as human disturbance. For example, two kittiwake colonies in Scotland (Dunbar Coast and the Isle of May) only 40km apart showed very different population trajectories, one increasing by 17% the other showing a moderate decline (Searle et al. 2023). Predation of both chicks and adult seabirds can impact breeding success and population size, e.g., by great skua or white-tailed eagle (Anker-Nilssen et al. 2023; Votier et al. 2004). Incidental seabird bycatch in fisheries can also impact seabirds, although this varies by species, fisheries gear and region, with fulmar bycatch in long-line fisheries a particular concern within Scotland (Northridge, Kingston & Coram, 2020).
4.9 Potential benefits to seabirds from sandeel fishery closure
The evidence above shows negative relationship between presence of a sandeel fishery and kittiwake breeding success, but limited evidence of a negative relationship for the other seabird species studied. However, Furness (2002) points out that the period of largest sandeel landings corresponds to the period of maximum seabird population size and breeding success over the last five decades. For example, UK kittiwake breeding numbers increased by 24% during 1969-1988 during which period the North Sea sandeel fishery greatly increased from landing less than 200 thousand tonnes annually to regularly landing around 800 thousand tonnes from 1977 onwards (JNCC 2021; Furness 2002). Despite the fishery removing large numbers of sandeel, seabird populations thrived. Furness (2002) hypothesised that reduced numbers of piscivorous fish had allowed a large increase in the sandeel stock that could sustain both seabirds and a large industrial fishery (Sherman et al, 1981). Furness (2002) proposed that the sandeel stock was regulated by bottom-up processes and so fishing and natural predation served to reduce density dependence in the stock and increased population growth. However, longer term, the stock did not continue to support high numbers of breeding seabirds. Whilst the fishery continued to land similar numbers of sandeel until 2000, kittiwake numbers declined in the UK by 25% between the Seabird Colony Register (1985-88) and Seabird 2000 (1998-2002) censuses, and subsequently sustained a further 29% decline by 2019 (JNCC 2021). Frederiksen et al. (2007) suggested that mature herring feeding on sandeel could be outcompeting seabirds around Shetland, observing a negative correlation between herring spawning stock biomass and sandeel total stock biomass. It is therefore possible that over this period piscivorous fish stocks have increased to the level that they may be outcompeting seabirds.
4.10 Role of environment
The impacts of industrial sandeel fishing and subsequent benefits following closure of the Wee Bankie fishery to seabirds during the breeding season has been extensively investigated (Daunt et al. 2008, Rindorf et al. 2000, Searle et al. 2023, Frederiksen et al. 2008, Wanless et al. 1998, Frederiksen et al. 2004, Cook et al. 2014, Wanless et al. 2004, Frederiksen et al. 2006, Frederiksen et al. 2005, Wanless et al. 2018). Whilst all studies identified some negative effect of the fishery and/or subsequent benefit following the closure, particularly on kittiwake, effects of the fishery on seabirds were complex and the role of environment on driving sandeel availability was also likely a dominant factor. The effects of the sandeel fishery may be additive on top of wider environmental processes, particularly climate change, that are reducing sandeel availability to seabirds (Rindorf et al. 2000). Therefore, closing the fishery may be beneficial but environmental processes will more strongly determine seabird breeding success (Cook et al. 2014, Daunt et al. 2008).
4.11 Seabird foraging ranges and existing sandeel fishery closed area
During the breeding season, seabirds are constrained to foraging within a certain distance from their breeding site due to the need to return to their nest to incubate eggs, feed chicks, brood young and protect eggs and chicks from predation. For seabirds to achieve improved breeding success from an increase in sandeel biomass from a fishery closure, the sandeel need to be within foraging range of colonies, in the water column (i.e., not buried in the sediment) and peak age 0 and age 1+ availability needs to coincide with the key phases of the breeding season.
Kittiwake have a mean foraging range of 55km and a mean maximum foraging range of 156km (Woodward et al. 2019). For many seabird colonies along the east coast of Scotland, this means that a typical foraging range would not regularly include foraging outside of the existing closed area (Figure 29). Whilst kittiwake are capable of flying further to the wider SA4 area, there would need to be considerably improved foraging opportunities above what is available in the existing closed area to make the longer foraging distance worthwhile. There is evidence that kittiwake forage at greater distances from their colonies when breeding success is low, suggesting birds will travel further when prey is not available locally (Hamer et al. 1993; Wanless et al. 1992). Unless sandeel metapopulation dynamics mean a wider sandeel closure would significantly change sandeel availability within the existing closed area, improved sandeel availability may generally be of limited benefit to kittiwake breeding success (see sections 3.1.2.2 and 3.1.2.3).
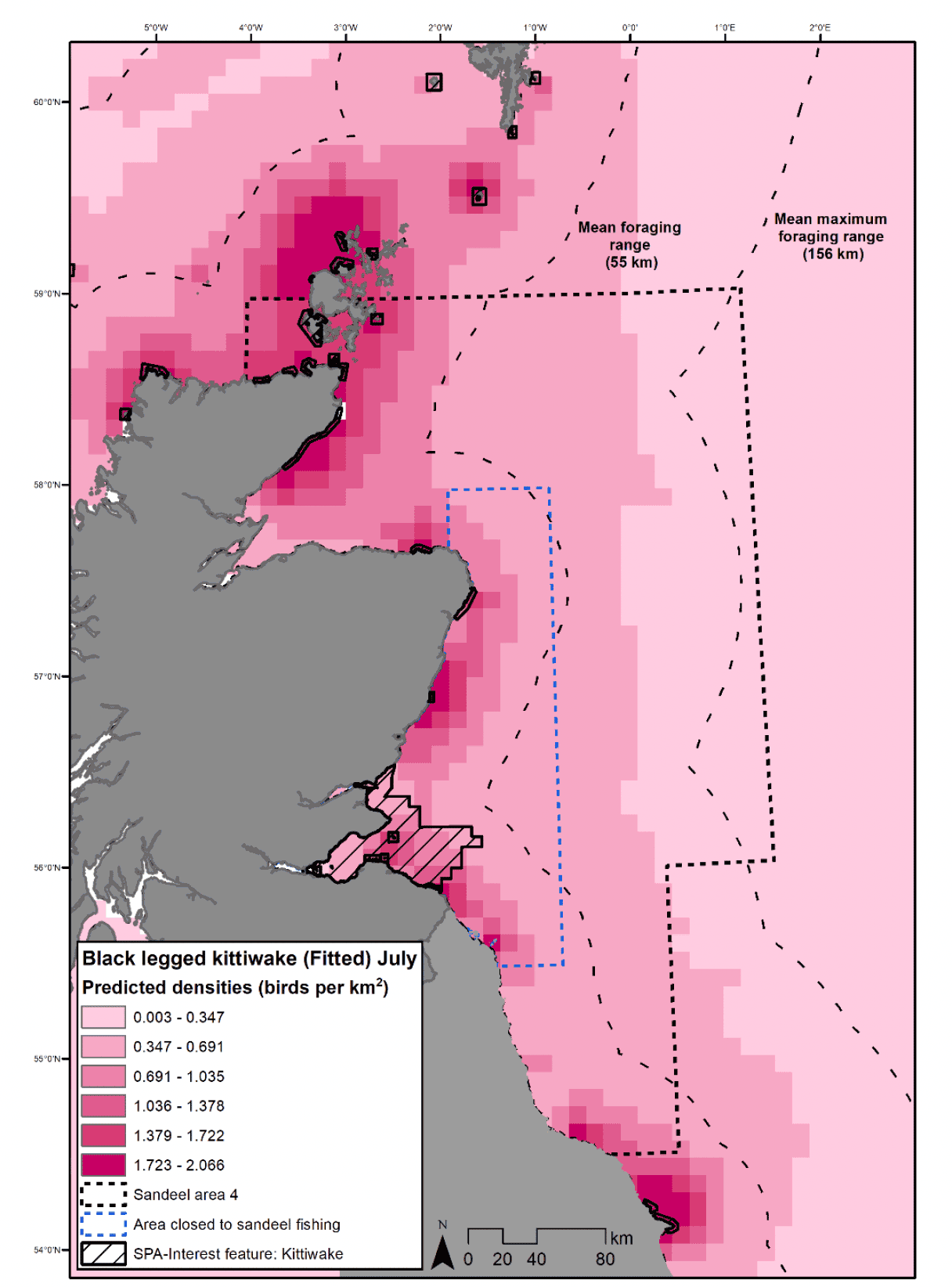
Some other seabirds have greater foraging ranges and could benefit from increases in sandeel availability, e.g., gannet with a mean foraging range of 120km, though this species is considered to have a low sensitivity to changes in sandeel availability. However, most species either have a similar mean foraging range to kittiwake, e.g., 61km and 62km for razorbill and puffin respectively, or they have a smaller mean foraging range than kittiwakes (Woodward et al. 2019). This means that for colonies within the existing area closed to sandeel fishing, with the exception of gannet, benefits of extending the closure will only be seen if either seabirds increase their mean foraging ranges and/or the closure changes availability of sandeel within the inshore existing closed area.
For species with colonies outside of the existing closed area, e.g., Orkney, Shetland, Caithness and Moray coast, benefits from a wider closure would be within their typical foraging ranges (Figure 29). However, as discussed above, these areas are not currently targeted by the sandeel fishery and benefits of a closure may be minimal.
While seabirds are not constrained to feeding around their colonies or provisioning offspring during the non-breeding period, the winter months are energetically challenging for many seabirds due to more inclement weather, limited daylight and reduced prey availability (Daunt et al. 2014). For example, winter sandeel spawning may provide an important prey resource prior to breeding allowing individuals to attain breeding condition. In addition, for species which can extract sandeel from the seabed, sandeel may provide an important prey throughout the non-breeding period, albeit at lower numbers/frequency compared to breeding (Howells et al. 2018). Thus, while the limited available evidence on seabird diet in the non-breeding period suggests that sandeel are less important relative to breeding, maintenance of sandeel stocks may confer some benefits to and resilience in seabird populations.
4.12 Variation in response to a sandeel fishery closure across seabird species
The extent to which seabirds might benefit from a wider sandeel fishery closure will vary among species. Benefits from increased sandeel abundance would be most likely to be seen in species that are most sensitive to changes in abundance, e.g., kittiwake and least in those species that can dive (guillemot, razorbill, puffin), switch to alternative prey (guillemot, gannet, great skua) or forage over a wider area (gannet) (Furness & Tasker, 2000).
Some seabird species have increased the proportion of clupeids (mostly sprat but some herring) in their diet in response to decreased sandeel availability (Wanless et al. 2018). Chick diet of kittiwake, guillemot, razorbill and puffin is almost entirely sandeel or sprat. This diet switch has helped buffer some species, e.g., guillemot, from decreases in sandeel abundance, compared with kittiwake. However, if sprat abundance also declines, these seabird species could see large reductions in breeding success.
Both Daunt et al. (2008) and Searle et al. (2023) did not detect any increase in breeding success following the Wee Bankie sandeel closure for any species other than kittiwake. Daunt et al. (2008) concluded this was because some species feed close inshore in unfished areas (terns, shag) or can dive in the water column (guillemot, razorbill and puffin) and so are less affected by a decrease in absolute abundance of sandeel than surface feeders. However, identifying or quantifying benefits of a sandeel fishery closure to seabirds, particularly with ongoing environmental change, is challenging (Rindorf et al. 2000, Searle et al. 2023, Daunt et al. 2008, Furness et al. 2007).
Seabird demography is also driven by variables besides prey availability, but increased prey availability might reduce the impact of other drivers, such as predation. Anker Nilssen et al. (2023) found that kittiwake breeding success and population size declined at a colony subject to white-tailed eagle predation, but that good foraging conditions for kittiwake slowed the rate of decline.
Whilst seabirds may not exhibit increases in breeding success because of the closure of Scottish waters to industrial sandeel fishing, there may be ecological benefits to seabirds which are more difficult to measure and quantify. For example, as described above, the larger sandeel stock will comprise a greater range of ages and sizes, which might increase the duration of sandeel availability to seabirds, fecundity, or population resilience to changing environmental conditions. This could in turn result in less extreme fluctuations in seabird breeding success and ensure greater resilience in kittiwake populations.
4.13 Summary of Evidence on Seabirds and Sandeel
Scotland supports internationally important populations of breeding seabirds, with 24 species regularly breeding in Scotland. Breeding seabirds are constrained in the distances from the nest site that they can forage, potentially limiting access to available sandeel and other prey resources. A large proportion of seabird species in Scotland include sandeel in their diet during the breeding season, though this dependence varies across species with gannet identified as having low sensitivity to sandeel abundance and kittiwake high sensitivity.
Kittiwake tend to feed on age 1+ sandeel in April and May, shifting to age 0 sandeel in June and July, with highest breeding success occurring when age 0 sandeel appeared early in the season. To be of most benefit to seabirds, the peak in sandeel abundance needs to coincide with the seabird chick rearing phase of the seabird breeding season. However, the timing of sandeel availability and absolute abundance shows inter-annual variation, which can result in a mismatch between peak sandeel availability and seabird chick rearing and negative effects on seabird productivity. Some seabird species are more able to switch prey species than others, for example guillemot switch prey species more easily than kittiwake, which may help mitigate some of the negative effects of reduced or mismatched sandeel availability.
Seabird breeding success has been shown to have a non-linear relationship with forage fish biomass, with breeding success rapidly declining as stocks fall below one third of maximum. Positive relationships between seabird breeding success and changes in forage fish abundance have been described for a range of seabird species.
Prey availability, rather than abundance or biomass, plays a key role in the breeding success of some seabirds. Prey need to be within foraging distance of seabird colonies, they need to be within the water column, and they need to be within dive depth (which varies considerably among seabird species). Similarly, prey of the right age or size class must be available at the right time of year for provisioning to chicks.
Establishing a relationship between industrial sandeel fisheries and seabird demography is extremely challenging. Only correlative relationships can be established meaning confidence in results can be low, and lag effects between seabird demography and environmental conditions increases complexity and uncertainty. Fishing mortality is only one factor influencing sandeel stock biomass, with natural predation by other fish, marine mammals, and seabirds, copepod prey abundance, and wider environmental conditions key factors.
On two occasions sandeel fisheries have been closed due to concerns about their impacts on the breeding success of seabirds: the small sandeel fishery off Shetland in the 1990s and the Wee Bankie, Scalp Bank and Marr Bank fishery in 2000. Searle et al. (2023) examined changes in breeding success and diet of kittiwake, guillemot, razorbill, and puffin during 1986-2018 in relation to the opening and closure of the Wee Bankie fishery. Kittiwake showed a significant decrease in breeding success attributable to the sandeel fishery (from 52% to 23% proportion of nests to young fledged), followed by a non-significant increase (10%) when the fishery closed, compared with control colonies. A key finding was that closing the fishery did not enable kittiwake breeding success to recover to pre-fishery levels, perhaps suggesting that wider environmental drivers were the dominant factors. Puffin, razorbill, and guillemot showed no negative effects of the fishery on breeding success, nor any positive effects following closure.
Over recent decades, the availability, size, and calorific content of sandeel has declined and some seabirds appear to have reduced their dependency on sandeel. Compensating for these reductions by increasing the number of prey items may be possible for some seabird species or populations. Therefore, whilst closing a sandeel fishery may be beneficial, wider environmental conditions are likely to determine seabird breeding success more strongly.
The positive benefits to seabird productivity and populations of a sandeel fishery closure are difficult to quantify because of the complex relationships between prey and seabird demography, ongoing climate-mediated changes in sandeel population (including from climate change) and the numerous other pressures that seabirds face. There is also considerable variation across seabird species in their dependence upon sandeel and their ability to switch to alternative prey. However, despite these uncertainties, maximising abundance and availability of sandeel stocks as prey for seabirds in Scotland remains a key mechanism by which resilience in seabird populations might be achieved.
Contact
Email: sandeelconsultation@gov.scot
There is a problem
Thanks for your feedback